Счет 75 «Расчеты с учредителями» / КонсультантПлюс
Счет 75 «Расчеты с учредителями»
Счет 75 «Расчеты с учредителями» предназначен для обобщения информации о всех видах расчетов с учредителями предприятия (акционерами акционерного общества, участниками хозяйственного товарищества, членами кооператива и т.п.): по вкладам в уставный фонд предприятия, по выплате доходов и др.
К счету 75 «Расчеты с учредителями» открывают субсчета:
75-1 «Расчеты по вкладам в уставный фонд»;
75-2 «Расчеты по доходам».
На субсчете 75-1 «Расчеты по вкладам в уставный фонд» учитывают расчеты с учредителями предприятия по вкладам в его уставный фонд.
При создании акционерного общества по дебету счета 75 «Расчеты с учредителями» в корреспонденции со счетом 85 «Уставный фонд» принимается на учет сумма задолженности лиц, подписавшихся на акции.
При фактическом поступлении сумм вкладов учредителей производят записи по кредиту счета 75 «Расчеты с учредителями» в корреспонденции со счетами по учету денежных средств.
При предоставлении зданий, сооружений и оборудования в пользование предприятию делают записи по кредиту счета 75 «Расчеты с учредителями» в корреспонденции со счетом 04 «Нематериальные активы». Одновременно на забалансовый учет по счету 001 «Арендованные основные средства» принимается стоимость указанных зданий, сооружений и оборудования.
Оприходование имущества, предоставленного в натуральной форме в собственность предприятия в счет вкладов в уставный фонд (в оплату акций), производят в оценке, определенной по договоренности участников. Оприходование имущества, предоставленного в натуральной форме в пользование предприятию в счет вкладов в уставный фонд (в оплату акций), производят в оценке, определенной исходя из арендной платы за пользование этим имуществом, исчисленной за весь указанный в учредительных документах срок, если иное не предусмотрено учредительными документами.
В аналогичном порядке отражают в бухгалтерском учете расчеты по вкладам в уставный фонд с участниками хозяйственных товариществ других видов. При этом запись по дебету счета 75 и кредиту счета 85 «Уставный фонд» производят на всю величину уставного фонда, объявленную в учредительных документах.
В том случае, когда акции предприятия, созданного в форме акционерного общества, реализуют по цене, превышающей номинальную стоимость их, вырученную сумму разницы между продажной и номинальной стоимостью относят в кредит счета 86 «Резервный фонд».
На субсчете 75-2 «Расчеты по доходам» учитывают расчеты с учредителями предприятия по выплате им доходов. Начисление и выплату доходов работникам предприятия, входящим в число его учредителей, учитывают на счете 70 «Расчеты с персоналом по оплате труда».
Начисление доходов от участия в предприятии отражают записью по дебету счета 87 «Нераспределенная прибыль (непокрытый убыток)» (за счет прибыли отчетного года или нераспределенной прибыли прошлых лет) и кредиту счета 75 «Расчеты с учредителями». При отсутствии или недостаточности прибыли для выплаты доходов, но при наличии обязательств, предусмотренных законодательством или учредительными документами, по выплате доходов за счет резервного фонда начисление доходов отражают по дебету счета 86 «Резервный фонд» и кредиту счета 75 «Расчеты с учредителями».
При отсутствии или недостаточности прибыли для выплаты доходов, но при наличии обязательств, предусмотренных законодательством или учредительными документами, по выплате доходов за счет резервного фонда начисление доходов отражают по дебету счета 86 «Резервный фонд» и кредиту счета 75 «Расчеты с учредителями».
Выплату начисленных сумм доходов отражают по дебету счета 75 «Расчеты с учредителями» в корреспонденции со счетом 51 «Расчетный счет или 52 «Валютный счет». При выплате доходов от участия в предприятии продукцией (работами, услугами) этого предприятия в бухгалтерском учете производят записи по дебету счета 75 «Расчеты с учредителями» в корреспонденции со счетом 46 «Реализация продукции (работ, услуг)».
Суммы налога на доходы от участия в предприятии, подлежащие уплате у источника, т.е. на этом предприятии, учитывают по дебету счета 75 «Расчеты с учредителями» и кредиту счета 68 «Расчеты с бюджетом».
Аналитический учет по счету 75 «Расчеты с учредителями» ведут по каждому учредителю, кроме учета расчетов с акционерами-собственниками акций на предъявителя и акционерными обществами.
Счет 75 «Расчеты с учредителями» корреспондирует со счетами:
┌──────────────────────────────────────────────────────┬─────────┐
│ Хозяйственная операция │Коррес- │
│ │пондирую-│
│ │щий счет │
├──────────────────────────────────────────────────────┼─────────┤
│ По дебету счета │ │
│ │ │
│Оплата начисленных дивидендов и иных обязательств │ 50, 51, │
│учредителей │ 52, 55 │
│ │ │
│Начисление налога на доходы от участия в предприятии │ 68 │
│ │ │
│Сумма задолженности лиц, подписавшихся на акции │ 85 │
│ │ │
│Расчеты по вкладам в уставный фонд │ 85 │
│ │ │
│Зачисление средств в резервный фонд │ 86 │
│ │ │
│Погашение непокрытого убытка отчетного года (прошлых │ 87 │
│лет) за счет целевых взносов учредителей (участников) │ │
│предприятия │ │
│ │ │
│Образование фондов специального назначения (накопления│ 88 │
│и потребления) за счет целевых взносов учредителей │ │
│(участников) предприятия │ │
│ │ │
│ По кредиту счета │ │
│ │ │
│Предоставление учредителями материальных и иных │ 01, 04, │
│ценностей (кроме денежных средств) в собственность │ 07, 08, │
│предприятия в счет взносов │ 10, 11, │
│ │ 12, 41 │
│ │ │
│Отражены предоставленные учредителями долгосрочные │ 06 │
│финансовые вложения в ценные бумаги, процентные │ │
│облигации государственных и местных займов, а также │ │
│представленные учредителями (участниками) другие │ │
│долгосрочные финансовые вложения │ │
│ │ │
│Поступление сумм вкладов учредителей в оплату │ 50, 51, │
│обязательств │ 52, 70 │
│ │ │
│Поступление ценных бумаг в оплату подписки на акции │ 58 │
│ │ │
│Начисление дивидендов акционерам (учредителям) за счет│ 81, 87 │
│прибыли, остающейся в распоряжении предприятия │ │
│ │ │
│Изменение уставного фонда за счет возврата вклада │ 85 │
│учредителю или уменьшения номинальной стоимости акций │ │
│ │ │
│Начисление дивидендов участникам за счет резервного │ 86 │
│фонда (при отсутствии или недостаточности прибыли │ │
│отчетного года для этих целей) │ │
│ │ │
│Распределение сумм прибыли, числящихся в фондах │ 88 │
│накопления, между учредителями предприятия │ │
└──────────────────────────────────────────────────────┴─────────┘
Учет расчетов с учредителями ООО, типовые проводки по счету 75
10 апреля 2014 Уставный капиталУчредителем ООО может выступать юридическое, а также физическое лицо РФ, иностранные организации и граждане. Действия при регистрации ООО и характер документации могут меняться в зависимости от состава участников общества. В статье разберем права и обязанности учредителей, учет взносов в уставный капитал и учет дивидендов, рассмотрим типовые проводки, связанные со взаиморасчетами с участниками организации.
Действия при регистрации ООО и характер документации могут меняться в зависимости от состава участников общества. В статье разберем права и обязанности учредителей, учет взносов в уставный капитал и учет дивидендов, рассмотрим типовые проводки, связанные со взаиморасчетами с участниками организации.
В общем, количество учредителей ООО не должно быть более 50 человек. Ими могут стать граждане, достигшие совершеннолетия, являющиеся дееспособными; ограничено дееспособные лица также имеют возможность заниматься предпринимательством, но только по согласию попечителя. Участники ООО не несут ответственности по его обязательствам, но несут риск получения убытков, которые связаны с деятельностью Общества и находятся в пределах стоимости долей, которые принадлежат им, согласно уставному капиталу ООО.
В процессе функционирования организации может потребоваться замена учредителя, как грамотно оформить эту процедуру читайте по ссылке.
Права и обязанности учредителей ООО
После регистрации у участников общества появляются определенные обязанности и права. Как гласит действующее законодательство РФ, участники ООО вправе:
Как гласит действующее законодательство РФ, участники ООО вправе:
- принимать участие в процессах управления делами ООО;
- иметь доступ к сведеньям о деятельности ООО и возможность ознакомления с его документацией в порядке, установленном уставом Общества;
- участвовать в процессе распределения прибыли;
- осуществить выход из ООО путем отчуждения доли, принадлежащей учредителю, Обществу, если это предусмотрено уставом ООО или стребовать приобретение доли Обществом;
- в случае ликвидации Общества получить часть имущества, которое осталось после расчета с кредиторами, либо получить стоимость этого имущества.
Кроме прав, учредители также имеют определенные обязанности. В их числе обязанность оплаты собственной доли в уставном капитале Общества и запрет на разглашение конфиденциальной информации, касающейся деятельности ООО. Участники ООО, которые не полностью оплатили долги, несут также солидарную ответственность. Она находится в пределах стоимости той части принадлежащих им долей, которая является не оплаченной.
Законом РФ может также быть запрещена или же ограничена возможность участия отдельных категорий граждан в ООО – к примеру, военные, муниципальные служащие не имеют права заниматься предпринимательством, в том числе являться учредителями коммерческих организаций.
Обязанности и права участников ООО физических лиц касаются и юридических лиц, но тут существуют некоторые ограничения. Согласно законодательству, органы местного управления и государственные органы имеют право являться учредителями ООО, если это не запрещено законом РФ.
Ответственность, которую несут участники ООО, является минимальной. Именно по этой причине общество с ограниченной ответственностью – самая распространенная организационно-правовая форма юридических лиц.
Бухгалтерский учет расчетов (проводки)
После того, как пройдена процедура регистрации ООО, помимо ведения предпринимательской деятельности появляется необходимость вести бухгалтерский учет на фирме.
Уставным капиталом называют сумму средств, которую готовы внести учредители в самом начале развития фирмы. В дальнейшем эти средства будут использоваться для деятельности предприятия. Подробнее об учете уставного капитала читайте здесь, а в этой статье подробно рассмотрен процесс изменения уставного капитала (увеличения и уменьшения).
В дальнейшем эти средства будут использоваться для деятельности предприятия. Подробнее об учете уставного капитала читайте здесь, а в этой статье подробно рассмотрен процесс изменения уставного капитала (увеличения и уменьшения).
Эти средства являют собой пассив предприятия, и будут являться источником формирования активов. Про активы и пассивы предприятия подробно говорили в этой статье.
Чтобы узнать, на каком счете рассчитывается уставной капитал, нужно выбрать его из Плана счетов. Есть сч. 80 «Уставной капитал». Его предназначение – учитывать этот капитал. Так как уставной капитал — пассив, значит, и сч. 80 тоже пассивный. Увеличение пассива отражается по кредиту, а уменьшение – дебету.
В Плане счетов также есть сч. 75 «Расчеты с учредителями». Этот счет нужен для осуществления всех расчетов с учредителями, то есть выплата дивидендов, взносы в уставной капитал и прочее.
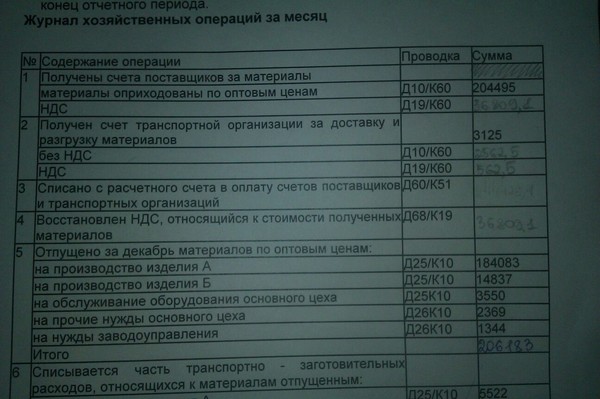
Проводки по расчетам с учредителями
По дебету 75 отражается величина уставного капитала (задолженность учредителей по взносам в уставный капитал).
По кредиту 75 отражается внесение учредителями взносов в уставный капитал.
Итак, составим проводки:
- Д75 К80 — отражена задолженность учредителей ООО по взносам в уставной капитал.
При погашении доли в уставном капитале задолженность учредителей перед организацией уменьшается. Это уменьшение будет отражаться по кредиту сч. 75. Второй счет, участвующий в проводке, выбирается в зависимости от вида взноса.
Взносы учредителей ООО в уставной капитал:
Если доля вносится безналичными средствами, вторым станет сч. 51 «Расчетный счет». По его дебету отразим взнос, поступающий от учредителя.
- Д51 К75 — взнос денежных средств (безналичных) на расчетный счет.
При внесении наличных средств, заносим сумму взноса в дебет сч. 50 «Касса».
- Д50 К75 — взнос в уставной капитал наличными.
Взнос также может осуществляться имуществом. При этом в качестве корреспондирующих счетов будут выступать счета: 10 «Материалы», 01 «Основные средства», 04 «Нематериальные активы», 41 «Товары» и т. д.
д.
Проводки по погашению учредителем доли в уставном капитале имуществом:
- Д10 К75 – проводка по отражению взноса в виде материалов;
- Д41 К75 — взнос в виде товаров и т.д.
Получение дивидендов учредителем:
Кроме взносов, на счете 75 также учитываются и дивиденды. Сначала происходит начисление дивидендов, после чего их выплата. Это две разные операции и отражаются они двумя проводками:
- Д84 К75 – проводка по начислению дивидендов учредителям из средств нераспределенной прибыли;
- Д75 К51 – проводка по выплате дивидендов учредителям с расчетного счета.
Типовые бухгалтерские проводки по учету расчетов с учредителями:
В процесс деятельности предприятия учредителем может быть оказана финансовая помощь, как на безвозмездной основе, так и возвратная. Подробнее об этом читайте здесь.
Счет 75 Расчеты с учредителями Раздела 6.
 Плана счетов бухгалтерского учета РБ
Плана счетов бухгалтерского учета РБСчет 75 «Расчеты с учредителями» предназначен для обобщения информации о расчетах с собственником имущества (учредителями, участниками) организации по вкладам в уставный фонд организации, выплате дивидендов и других доходов от участия в уставном фонде организации и др.
(в ред. постановления Минфина от 20.12.2012 N 77)
При создании акционерных обществ дебиторская задолженность учредителей по оплате акций отражается по дебету счета 75 «Расчеты с учредителями» и кредиту счета 80 «Уставный капитал». Суммы внесенных учредителями вкладов в погашение дебиторской задолженности по оплате акций отражаются по дебету счетов 50 «Касса», 51 «Расчетные счета», 52 «Валютные счета» и других счетов и кредиту счета 75 «Расчеты с учредителями». Если акции акционерного общества реализуются по цене, превышающей их номинальную стоимость, разница между ценой реализации акций и их номинальной стоимостью отражается на счете 83 «Добавочный капитал».
При создании организаций (за исключением акционерных обществ) дебиторская задолженность учредителей (участников) по вкладам в уставный фонд отражается по дебету счета 75 «Расчеты с учредителями» и кредиту счета 80 «Уставный капитал».
Изменение собственника имущества (учредителей, участников) или изменение величины его (их) вкладов (долей, паев) в уставном фонде отражаются внутренними записями по счету 75 «Расчеты с учредителями».
Увеличение уставного фонда отражается по дебету счета 75 «Расчеты с учредителями» и кредиту счета 80 «Уставный капитал».
(в ред. постановления Минфина от 20.12.2012 N 77)
Начисление дивидендов и других доходов от участия в уставном фонде организации отражается по дебету счета 84 «Нераспределенная прибыль (непокрытый убыток)» и кредиту счета 75 «Расчеты с учредителями». Выплата дивидендов и других доходов от участия в уставном фонде организации отражается по дебету счета 75 «Расчеты с учредителями» и кредиту счетов 50 «Касса», 51 «Расчетные счета», 52 «Валютные счета». Удержание налога на доходы у источника выплаты доходов отражается по дебету счета 75 «Расчеты с учредителями» и кредиту счета 68 «Расчеты по налогам и сборам».
Удержание налога на доходы у источника выплаты доходов отражается по дебету счета 75 «Расчеты с учредителями» и кредиту счета 68 «Расчеты по налогам и сборам».
Реструктуризация кредиторской задолженности в соответствии с законодательством отражается по дебету счетов 60 «Расчеты с поставщиками и подрядчиками», 66 «Расчеты по краткосрочным кредитам и займам», 67 «Расчеты по долгосрочным кредитам и займам», 68 «Расчеты по налогам и сборам», 69 «Расчеты по социальному страхованию и обеспечению», 76 «Расчеты с разными дебиторами и кредиторами» и других счетов и кредиту счета 75 «Расчеты с учредителями».
Аналитический учет по счету 75 «Расчеты с учредителями» ведется по каждому учредителю, участнику.
Счет 75 «Расчеты с учредителями» имеет следующие субсчета:
Счет 75 «Расчеты с учредителями» корреспондирует со счетами:
Другие счета раздела 6
Счет 60. Расчеты с поставщиками и подрядчиками Счет 62. Расчеты с покупателями и заказчиками Счет 63.
 Резервы по сомнительным долгам
Счет 65. Отложенные налоговые обязательства
Счет 66. Расчеты по краткосрочным кредитам и займам
Счет 67. Расчеты по долгосрочным кредитам и займам
Счет 68. Расчеты по налогам и сборам
Счет 69. Расчеты по социальному страхованию и обеспечению
Счет 70. Расчеты с персоналом по оплате труда
Счет 71. Расчеты с подотчетными лицами
Счет 73. Расчеты с персоналом по прочим операциям
Счет 75. Расчеты с учредителями
Счет 76. Расчеты с разными дебиторами и кредиторами
Счет 77. Расчеты по прямому страхованию и перестрахованию
Счет 79. Внутрихозяйственные расчеты
Резервы по сомнительным долгам
Счет 65. Отложенные налоговые обязательства
Счет 66. Расчеты по краткосрочным кредитам и займам
Счет 67. Расчеты по долгосрочным кредитам и займам
Счет 68. Расчеты по налогам и сборам
Счет 69. Расчеты по социальному страхованию и обеспечению
Счет 70. Расчеты с персоналом по оплате труда
Счет 71. Расчеты с подотчетными лицами
Счет 73. Расчеты с персоналом по прочим операциям
Счет 75. Расчеты с учредителями
Счет 76. Расчеты с разными дебиторами и кредиторами
Счет 77. Расчеты по прямому страхованию и перестрахованию
Счет 79. Внутрихозяйственные расчеты%PDF-1.7 % 830 0 объект > эндообъект внешняя ссылка 830 104 0000000016 00000 н 0000003964 00000 н 0000004140 00000 н 0000004176 00000 н 0000004367 00000 н 0000004592 00000 н 0000005259 00000 н 0000005362 00000 н 0000005630 00000 н 0000008655 00000 н 0000009052 00000 н 0000009460 00000 н 0000010065 00000 н 0000010579 00000 н 0000011122 00000 н 0000011290 00000 н 0000011943 00000 н 0000012201 00000 н 0000015701 00000 н 0000016108 00000 н 0000016478 00000 н 0000017072 00000 н 0000017128 00000 н 0000017516 00000 н 0000017723 00000 н 0000018010 00000 н 0000018639 00000 н 0000019215 00000 н 0000019750 00000 н 0000019999 00000 н 0000023254 00000 н 0000023636 00000 н 0000024017 00000 н 0000024677 00000 н 0000025386 00000 н 0000025766 00000 н 0000026186 00000 н 0000034095 00000 н 0000034618 00000 н 0000034728 00000 н 0000155907 00000 н 0000282295 00000 н 0000396245 00000 н 0000517148 00000 н 0000517206 00000 н 0000517305 00000 н 0000517390 00000 н 0000517506 00000 н 0000517618 00000 н 0000517746 00000 н 0000517868 00000 н 0000517988 00000 н 0000518150 00000 н 0000518311 00000 н 0000518458 00000 н 0000518606 00000 н 0000518750 00000 н 0000518919 00000 н 0000519028 00000 н 0000519190 00000 н 0000519311 00000 н 0000519448 00000 н 0000519630 00000 н 0000519751 00000 н 0000519864 00000 н 0000520034 00000 н 0000520133 00000 н 0000520260 00000 н 0000520450 00000 н 0000520608 00000 н 0000520734 00000 н 0000520920 00000 н 0000521034 00000 н 0000521197 00000 н 0000521312 00000 н 0000521431 00000 н 0000521557 00000 н 0000521699 00000 н 0000521827 00000 н 0000521989 00000 н 0000522125 00000 н 0000522365 00000 н 0000522515 00000 н 0000522729 00000 н 0000522851 00000 н 0000522985 00000 н 0000523125 00000 н 0000523275 00000 н 0000523417 00000 н 0000523627 00000 н 0000523823 00000 н 0000524005 00000 н 0000524185 00000 н 0000524299 00000 н 0000524459 00000 н 0000524599 00000 н 0000524737 00000 н 0000524879 00000 н 0000525021 00000 н 0000525159 00000 н 0000525267 00000 н 0000525393 00000 н 0000525525 00000 н 0000002376 00000 н трейлер ]/предыдущая 3998512>> startxref 0 %%EOF 933 0 объект >поток hвязьVkPWل4f#C56PA1+B`hJ]P!@q:AYPE5hE3B;;s9ιw
%PDF-1. 6
%
764 0 объект
>/Метаданные 761 0 R/AcroForm 840 0 R/Страницы 685 0 R/Тип/Каталог/Ярлыки Страницы 681 0 R>>
эндообъект
761 0 объект
>поток
UUID: 9c4a2a16-d682-49fd-a659-3f26b697a7c7adobe: DocId: INDD: 4b1e7f42-64d5-11dd-9e37-89d8589d7214proof: pdf9dd70365-5fd5-11dd-9e41-b993d393ebceadobe: DocId: INDD: b037dc68-a735-11dc-9f96-fb81a1572da82008- 08-12T11:07:49-04:002008-08-12T17:01:12-04:002008-08-12T17:01:12-04:00Adobe InDesign CS3 (5.0.3)
6
%
764 0 объект
>/Метаданные 761 0 R/AcroForm 840 0 R/Страницы 685 0 R/Тип/Каталог/Ярлыки Страницы 681 0 R>>
эндообъект
761 0 объект
>поток
UUID: 9c4a2a16-d682-49fd-a659-3f26b697a7c7adobe: DocId: INDD: 4b1e7f42-64d5-11dd-9e37-89d8589d7214proof: pdf9dd70365-5fd5-11dd-9e41-b993d393ebceadobe: DocId: INDD: b037dc68-a735-11dc-9f96-fb81a1572da82008- 08-12T11:07:49-04:002008-08-12T17:01:12-04:002008-08-12T17:01:12-04:00Adobe InDesign CS3 (5.0.3)
 0Ложь конечный поток
эндообъект
840 0 объект
>/Кодировка>>>>>
эндообъект
685 0 объект
>
эндообъект
681 0 объект
>
эндообъект
839 0 объект
>
эндообъект
837 0 объект
>
эндообъект
834 0 объект
>
эндообъект
832 0 объект
0Ложь конечный поток
эндообъект
840 0 объект
>/Кодировка>>>>>
эндообъект
685 0 объект
>
эндообъект
681 0 объект
>
эндообъект
839 0 объект
>
эндообъект
837 0 объект
>
эндообъект
834 0 объект
>
эндообъект
832 0 объектЦепь памяти человека, полученная в результате поражения головного мозга, вызывающего амнезию

Папез, Дж. В. Предлагаемый механизм эмоций. Арх. Нейрол. Психиатрия 38 , 725–743 (1937).
Артикул Google Scholar
Scoville, W. B. & Milner, B. Потеря недавней памяти после двусторонних поражений гиппокампа. Дж. Нейрол. Нейрохирург. психиатрия 20 , 11 (1957).
КАС Статья Google Scholar
Фон Крамон, Д.Ю., Хебель, Н. и Шури, У. Вклад в анатомические основы таламической амнезии. Мозг 108 , 993–1008 (1985).
Артикул Google Scholar
Valenstein, E. et al. Ретроспленальная амнезия. Мозг 110 , 1631–1646 (1987).
Артикул Google Scholar
Коркин, С. Что нового в пациенте с амнезией Х. М.? Нац.Преподобный Нейроски. 3 , 153 (2002).
КАС Статья Google Scholar
Swanson, L. W. & Cowan, W. M. Гиппокампо-гипоталамические связи: происхождение в субикулярной коре, а не в аммоновом роге. Наука 189 , 303–304 (1975).
ОБЪЯВЛЕНИЕ КАС Статья Google Scholar
Зола-Морган, С., Сквайр, Л. Р.и Мишкин, М. Нейроанатомия амнезии: миндалевидное тело-гиппокамп против височного ствола. Наука 218 , 1337–1339 (1982).
ОБЪЯВЛЕНИЕ КАС Статья Google Scholar
Моррис Р., Пандья Д. Н. и Петридес М. Система волокон, связывающая среднедорсолатеральную лобную кору с ретроспленальной/пресубикулярной областью у макак-резусов. Дж. Комп. Нейрол. 407 , 183–192 (1999).
КАС Статья Google Scholar
Choo, I. H. et al. Взаимосвязь фибриллярного бета-амилоидного бремени с возрастом начала болезни Альцгеймера. Болезнь Альцгеймера. Демент. Дж. Болезнь Альцгеймера. доц. 6 , С39–С40 (2010 г.).
H. et al. Взаимосвязь фибриллярного бета-амилоидного бремени с возрастом начала болезни Альцгеймера. Болезнь Альцгеймера. Демент. Дж. Болезнь Альцгеймера. доц. 6 , С39–С40 (2010 г.).
Артикул Google Scholar
Vincent, J.L. et al. Когерентная спонтанная активность идентифицирует гиппокампально-теменную сеть памяти. J. Нейрофизиол. 96 , 3517–3531 (2006).
Артикул Google Scholar
Wang, L. et al. Изменения в связях гиппокампа на ранних стадиях болезни Альцгеймера: данные фМРТ в состоянии покоя. Neuroimage 31 , 496–504 (2006).
Артикул Google Scholar
Грейциус, М. Д., Шривастава, Г., Reiss, A.L. & Menon, V. Сетевая активность в режиме по умолчанию отличает болезнь Альцгеймера от здорового старения: данные функциональной МРТ. Проц. Натл акад. науч. США 101 , 4637–4642 (2004 г.).
Натл акад. науч. США 101 , 4637–4642 (2004 г.).
ОБЪЯВЛЕНИЕ КАС Статья Google Scholar
Sestieri, C., Corbetta, M., Romani, G.L. & Shulman, G.L. Извлечение эпизодической памяти, теменная кора и сеть режима по умолчанию: функциональный и топографический анализ. J. Neurosci. 31 , 4407–4420 (2011).
КАС Статья Google Scholar
Болезнь Альцгеймера, А. Факты и цифры о болезни Альцгеймера, 2015 г. Болезнь Альцгеймера. Демент. Дж. Болезнь Альцгеймера. доц. 11 , 332 (2015).
Артикул Google Scholar
Weuve, J., Hebert, L.E., Scherr, P.A. & Evans, D.A. Распространенность болезни Альцгеймера в штатах США. Эпидемиология 26 , e4–e6 (2015).
Артикул Google Scholar

Lanctôt, K.L. et al. Нейропсихиатрические признаки и симптомы болезни Альцгеймера: новые парадигмы лечения. Болезнь Альцгеймера. Демент. Перевод Рез. клин. Интерв. 3 , 440–449 (2017).
Артикул Google Scholar
Hamani, C. et al. Улучшение памяти, вызванное глубокой стимуляцией гипоталамуса/свода мозга. Анналы неврологии: официальный . Дж. Ам. Нейрол. доц. Детский Нейрол. соц. 63 , 119–123 (2008).
Google Scholar
Suthana, N. et al. Улучшение памяти и глубокая стимуляция энторинальной области мозга. Новый англ. Дж. Мед. 366 , 502–510 (2012).
КАС Статья Google Scholar
Ван, Дж.Х. и др. Целенаправленное усиление корково-гиппокампальных сетей мозга и ассоциативной памяти. Наука 345 , 1054–1057 (2014).
ОБЪЯВЛЕНИЕ КАС Статья Google Scholar
Koch, G. et al. Транскраниальная магнитная стимуляция предклинья улучшает память и нервную активность при продромальной стадии болезни Альцгеймера. Нейроизображение 169 , 302–311 (2018).
Артикул Google Scholar
Aggleton, J.P., Pralus, A., Nelson, A.J. & Hornberger, M. Таламическая патология и потеря памяти при ранней болезни Альцгеймера: перемещение фокуса из медиальной височной доли в контур Папеса. Мозг 139 , 1877–1890 (2016).
Артикул Google Scholar
Boes, A.D. et al. Сетевая локализация неврологической симптоматики при очаговых поражениях головного мозга. Мозг 138 , 3061–3075 (2015).
Артикул Google Scholar

Дарби, Р. Р., Хорн, А., Кушман, Ф. и Фокс, М. Д. Локализация сети повреждений преступного поведения. Проц. Натл акад. науч. США 115 , 601–606 (2018).
КАС Статья Google Scholar
Fox, M.D. Сопоставление симптомов с сетями мозга с помощью коннектома человека. Новый англ. Дж. Мед. 379 , 2237–2245 (2018).
КАС Статья Google Scholar
Corbetta, M. et al. Общие поведенческие кластеры и подкорковая анатомия при инсульте. Нейрон 85 , 927–941 (2015).
КАС Статья Google Scholar
Рэймонт, В., Салазар, А., Крюгер, Ф. и Графман, Дж. «Изучение психических травм» – исследование черепно-мозговых травм во Вьетнаме и 40 лет исследований черепно-мозговых травм. Фронт. Нейрол. 2 , 15 (2011).
Артикул Google Scholar
Fox, M.D. et al. Человеческий мозг внутренне организован в виде динамических, антикоррелированных функциональных сетей. Проц. Натл акад. науч. США 102 , 9673–9678 (2005 г.).
ОБЪЯВЛЕНИЕ КАС Статья Google Scholar
Yeo, B.T. et al.Организация коры головного мозга человека оценивается по внутренней функциональной связности. J. Нейрофизиол. 106 , 1125–1165 (2011).
Артикул Google Scholar
Raichle, M.E. et al. Режим работы мозга по умолчанию. Проц. Натл акад. науч. США 98 , 676–682 (2001).
ОБЪЯВЛЕНИЕ КАС Статья Google Scholar
Бакнер, Р. Л., Эндрюс-Ханна, Дж. Р. и Шактер, Д. Л. Сеть мозга по умолчанию. Энн. Н.Ю. акад. науч. 1124 , 1–38 (2008).
ОБЪЯВЛЕНИЕ Статья Google Scholar
Спренг, Р. Н. и Грейди, К. Л. Паттерны активности мозга, поддерживающие автобиографическую память, исследование и теорию разума, и их связь с сетью режима по умолчанию. Дж. Когн. Неврологи. 22 , 1112–1123 (2010).
Артикул Google Scholar
Рорден С., Карнат Х. О. и Бонилья Л. Улучшение картирования симптомов поражения. Дж. Когн. Неврологи. 19 , 1081–1088 (2007).
Артикул Google Scholar
Tulving, E. et al. Полушарная асимметрия кодирования/поиска в эпизодической памяти: данные позитронно-эмиссионной томографии. Проц. Натл акад.науч. США 91 , 2016–2020 (1994).
ОБЪЯВЛЕНИЕ КАС Статья Google Scholar
Shallice, T. et al. Области мозга, связанные с приобретением и воспроизведением вербальной эпизодической памяти. Природа 368 , 633 (1994).
ОБЪЯВЛЕНИЕ КАС Статья Google Scholar
Уилер, М. А., Стусс, Д. Т. и Талвинг, Э.К теории эпизодической памяти: лобные доли и автоноэтическое сознание. Психология. Бык. 121 , 331 (1997).
КАС Статья Google Scholar
Паккард, М. Г. и Ноултон, Б. Дж. Функции обучения и памяти базальных ганглиев. год. Преподобный Нейроски. 25 , 563–593 (2002).
КАС Статья Google Scholar
Вагнер, А. Д., Шеннон, Б. Дж., Кан, И. и Бакнер, Р. Л. Вклад теменной доли в поиск эпизодической памяти. Тенденции Cogn. науч. 9 , 445–453 (2005).
Артикул Google Scholar
Шмахманн Дж. Д. и Каплан Д. Познание, эмоции и мозжечок. Мозг 129 , 290–292 (2006).
Артикул Google Scholar
Sestieri, C. et al. Нарушение восстановления эпизодической памяти после транскраниальной стимуляции нижней, но не верхней теменной дольки. Нейропсихология 51 , 900–906 (2013).
Артикул Google Scholar
Лю Т. и др. Извилистость коры и борозды на ранней стадии болезни Альцгеймера. PloS ONE 7 , e31083 (2012).
ОБЪЯВЛЕНИЕ КАС Статья Google Scholar
Далтон, М. А. и Магуайр, Э. А. Пре/парасубикулум: гиппокампальный узел для познания на основе сцены? Курс. мнение Поведение науч. 17 , 34–40 (2017).
Артикул Google Scholar
Поппенк Дж., Эвенсмун Х.Р., Москович М. и Надель Л. Специализация гиппокампа человека по длинной оси. Тенденции Cogn.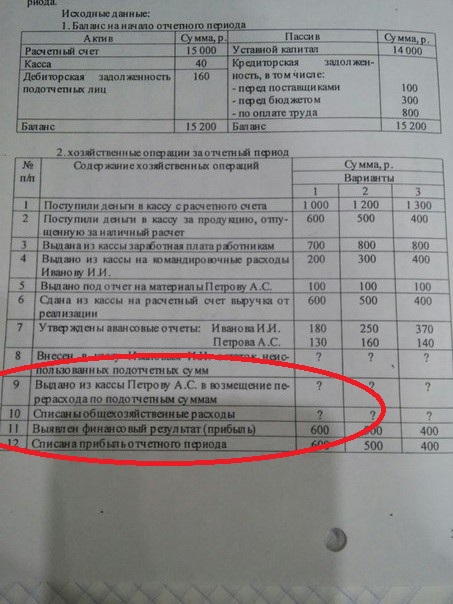 науч. 17 , 230–240 (2013).
науч. 17 , 230–240 (2013).
Артикул Google Scholar
Zeidman, P. & Maguire, EA Передний гиппокамп: анатомия восприятия, воображения и эпизодической памяти. Нац. Преподобный Нейроски. 17 , 173 (2016).
КАС Статья Google Scholar
Eichenbaum, H. Префронтально-гиппокампальные взаимодействия в эпизодической памяти. Нац. Преподобный Нейроски. 18 , 547 (2017).
КАС Статья Google Scholar
Роллс, Э. Т. Хранение и воспроизведение воспоминаний в гиппокамп-кортикальной системе. Клеточная ткань Рез. 373 , 577–604 (2018).
КАС Статья Google Scholar
Мацумото, Н., Китаниши, Т., и Мидзусеки, К. Субикулум: уникальный центр гиппокампа и многое другое. Неврологи. Рез. 143 , 1–12 (2018).
Неврологи. Рез. 143 , 1–12 (2018).
Roy, D.S. et al. Отдельные нейронные цепи для формирования и извлечения эпизодических воспоминаний. Ячейка 170 , 1000–1012 (2017).
КАС Статья Google Scholar
О’Мара, С. Субикулум: что он делает, что он может делать и что нам еще предстоит рассказать нейроанатомии. Дж. Анат. 207 , 271–282 (2005).
Артикул Google Scholar
Wyass, JM & Van Groen, T. Связи между ретроспленальной корой и образованием гиппокампа у крыс: обзор. Гиппокамп 2 , 1–11 (1992).
Артикул Google Scholar
Чайлд Н.Д. и Бенаррох Э.Е. Переднее ядро таламуса: функциональная организация и клинические последствия. Неврология 81 , 1869–1876 (2013).
Артикул Google Scholar
Вивиани Р. Цифровой атлас сосудов среднего и крупного мозга и их связь с корковыми и подкорковыми структурами. Фронт. Нейроанат. 10 , 12 (2016).
Артикул Google Scholar
Бакнер, Р. Л., Кринен, Ф. М. и Йео, Б. Т. Возможности и ограничения внутренней функциональной связи МРТ. Нац. Неврологи. 16 , 832 (2013).
Артикул Google Scholar
Cole, M.W. et al. Внутренние и вызванные задачами сетевые архитектуры человеческого мозга. Нейрон 83 , 238–251 (2014).
КАС Статья Google Scholar
Dosenbach, N.U. et al. Отдельные мозговые сети для адаптивного и стабильного управления задачами у людей. Проц. Натл акад. науч. США 104 , 11073–11078 (2007 г.).
ОБЪЯВЛЕНИЕ КАС Статья Google Scholar

Tavor, I. et al. Беззадачная МРТ предсказывает индивидуальные различия в активности мозга во время выполнения задачи. Наука 352 , 216–220 (2016).
ОБЪЯВЛЕНИЕ КАС Статья Google Scholar
Ито, Т. и др. Информация о когнитивных задачах передается между областями мозга через топологию сети в состоянии покоя. Нац. коммун. 8 , 1027 (2017).
ОБЪЯВЛЕНИЕ Статья Google Scholar
Cole, M.W. et al. Активация задачи приводит к ложному, но систематическому завышению оценок функциональной связности задачи. NeuroImage 189 , 1–18 (2019).
Артикул Google Scholar
Fox, M.D. & Raichle, M.E. Спонтанные колебания активности мозга, наблюдаемые с помощью функциональной магнитно-резонансной томографии. Нац. Преподобный Нейроски. 8 , 700 (2007 г.).
8 , 700 (2007 г.).
КАС Статья Google Scholar
Адольф Р. Исследования повреждений человека в 21 веке. Нейрон 90 , 1151–1153 (2016).
КАС Статья Google Scholar
Schroeter, M.L. & Neumann, J. Комбинированные визуализирующие маркеры диссоциируют болезнь Альцгеймера и дегенерацию лобно-височных долей – метаанализ ALE. Границы старения . Неврология 3 , 10 (2011).
Google Scholar
Darby, R., Dickerson, B.C. & Fox, M. Антикоррелированные сети показывают парадоксальное увеличение толщины коры при болезни Альцгеймера. Болезнь Альцгеймера. Демент. Дж. Болезнь Альцгеймера. доц. 14 , P834 (2018).
Артикул Google Scholar
Gultekin, S.H. et al. Паранеопластический лимбический энцефалит: неврологические симптомы, иммунологические данные и ассоциация с опухолью у 50 пациентов. Мозг 123 , 1481–1494 (2000).
et al. Паранеопластический лимбический энцефалит: неврологические симптомы, иммунологические данные и ассоциация с опухолью у 50 пациентов. Мозг 123 , 1481–1494 (2000).
Артикул Google Scholar
Bartsch, T. & Deuschl, G. Преходящая глобальная амнезия: функциональная анатомия и клинические последствия. Ланцет Нейрол. 9 , 205–214 (2010).
Артикул Google Scholar
Barash, J.A. et al. Острый амнестический синдром, связанный с передозировкой фентанила. Новый англ. Дж. Мед. 378 , 1157–1158 (2018).
Артикул Google Scholar
Fisher, D.B. et al. Сетевая неинвазивная стимуляция мозга мультифокальными tdcs. Стимуляция мозга. Базовый, пер., клин. Рез. Нейромодуляция 10 , 411–412 (2017).
Google Scholar

Holmes, A.J. et al. Выпуск исходных данных Brain Genomics Superstruct Project со структурными, функциональными и поведенческими показателями. Науч. Данные 2 , 150031 (2015 г.).
Артикул Google Scholar
Фокс, М. Д., Чжан, Д., Снайдер, А. З. и Райхл, М. Э. Глобальный сигнал и наблюдаемые антикоррелированные сети мозга в состоянии покоя. J. Нейрофизиол. 101 , 3270–3283 (2009 г.).
Артикул Google Scholar
Horn, A. et al. Connectivity Предсказывает результат глубокой стимуляции мозга при болезни Паркинсона. Энн. Нейрол. 82 , 67–78 (2017).
Артикул Google Scholar
Ву, О. и др. Роль топографии острого поражения в исходной тяжести ишемического инсульта и отдаленных функциональных исходах. Инсульт 46 , 2438–2444 (2015).
КАС Статья Google Scholar
Iglesias, J.E. et al. Вычислительный атлас образования гиппокампа с использованием ex vivo МРТ сверхвысокого разрешения: применение к адаптивной сегментации МРТ in vivo. Нейроизображение 115 , 117–137 (2015).
Артикул Google Scholar
Графман Дж., Салазар А. М., Вайнгартнер Х. и Амин Д. Память лица и различение: анализ стойких последствий проникающих ранений головного мозга. Междунар. Дж. Нейроски. 29 , 125–139 (1986).
Артикул Google Scholar
Альберт М.С., Баттерс Н. и Левин Дж. Временные градиенты при ретроградной амнезии у пациентов с алкогольной болезнью Корсакова. Арка Нейрол. 36 , 211–216 (1979).
КАС Статья Google Scholar

Бенуа, Р. Г. и Шактер, Д. Л. Определение базовой сети, поддерживающей эпизодическую симуляцию и эпизодическую память, путем оценки вероятности активации. Нейропсихология 75 , 450–457 (2015).
Артикул Google Scholar
Rottschy, C. et al. Моделирование нейронных коррелятов рабочей памяти: метаанализ на основе координат. Нейроизображение 60 , 830–846 (2012).
КАС Статья Google Scholar
Ferstl, E.C., Neumann, J., Bogler, C. & Von Cramon, D.Y. Расширенная языковая сеть: метаанализ нейровизуализационных исследований понимания текста. Гул. Карта мозга. 29 , 581–593 (2008).
Артикул Google Scholar
Schroeter, M.L., Raczka, K., Neumann, J. & Von Cramon, D.Y. Нейронные сети при лобно-височной деменции — метаанализ. Нейробиол. Старение 29 , 418–426 (2008).
Старение 29 , 418–426 (2008).
Артикул Google Scholar
Бисениус С., Нойманн Дж.& Schroeter, M.L. Валидация новых критериев диагностической визуализации для первичной прогрессирующей афазии с помощью метаанализа анатомической оценки правдоподобия. евро. Дж. Нейрол. 23 , 704–712 (2016).
КАС Статья Google Scholar
Биология нейронной регуляции
Колд-Спринг-Харб Perspect Biol. 2010 июнь; 2(6): а001917.
Alain Chédotal
1 INSERM UMRS_968, Institut de la Vision, Департамент развития, 17 rue Moreau, 75012 Париж, Франция; UPMC Univ Paris 06, UMRS_968, F-75012 Paris, France
Линда Дж.Richards
2 Университет Квинсленда, Квинслендский институт мозга и Школа биомедицинских наук, здание 79, кампус Сент-Люсия, Сент-Луика, Квинсленд, Австралия, 4072
1 INSERM UMRS_968, Институт зрения, Департамент Девелопмент, улица Моро, 17, 75012 Париж, Франция; UPMC Univ Paris 06, UMRS_968, F-75012 Paris, France
2 Университет Квинсленда, Квинслендский институт мозга и школа биомедицинских наук, здание 79, кампус Сент-Люсия, Сент-Луика, Квинсленд, Австралия, 4072
Copyright © 2010 Издательство лаборатории Колд-Спринг-Харбор; Все права защищены. Эта статья цитировалась в других статьях PMC.
Эта статья цитировалась в других статьях PMC.Abstract
Мозг млекопитающих является наиболее сложным органом в организме. Он контролирует все аспекты наших телесных функций и интерпретирует окружающий мир через наши чувства. Он определяет нас как людей через наши воспоминания и нашу способность планировать будущее. Решающее значение для всех этих функций имеет то, как мозг устроен для выполнения этих задач. Основная карта проводки мозга возникает во время эмбрионального и постнатального развития посредством серии точно организованных событий развития, регулируемых специфическими молекулярными механизмами.Ниже мы рассмотрим наиболее важные особенности проводки мозга млекопитающих, полученные в результате работы как у млекопитающих, так и у других видов. Эти механизмы сохранялись на протяжении всей эволюции, просто усложняясь в мозге млекопитающих. Эта захватывающая область биологии раскрывает суть того, что позволяет мозгу млекопитающих выполнять повседневные задачи, которые мы считаем само собой разумеющимися, а также те, которые дают нам возможность для выдающихся достижений.
СТРУКТУРА МОЗГА В СВЯЗИ С ПЕРВЫМИ АКСОНАМИ
В нервной системе взрослого человека аксоны, происходящие из групп нейронов в ядрах, определенных клеточных слоях или ганглиях, преимущественно ассоциируются, по крайней мере, на части своего пути, с четко определенным белым веществом тракты точно распределены вдоль рострокаудальной и дорсовентральной осей.Эта особенность объясняется предпочтительным ростом развивающихся аксонов вдоль ранее существовавших аксональных путей (Goodman and Shatz 1993). Тем не менее, первые аксоны, которые появляются в развивающемся мозге, растут в значительной степени без аксонов, проводя поверхностно через недифференцированные нейроэпителиальные клетки. Эти ранние аксоны называются «пионерами», и считается, что они прокладывают путь, по которому следуют более поздние растущие аксоны (Easter et al. 1994). Аксоны, прибывающие позже, имеют тенденцию объединяться с пионерами через установленный «каркас», который обеспечивает базовую основу для аксонов «последователей». Покадровые исследования показали, что морфология конусов роста, поведение и динамика актина аксонов-первопроходцев отличаются от таковых у аксонов-последователей, которые менее сложны, растут с большей скоростью через точки выбора и имеют более высокую динамику актина (Bak и Фрейзер, 2003; Кулкарни и др., 2007). Роль пионерных аксонов в направлении последующих (из одного ядра или другого происхождения) в первую очередь подтверждается экспериментами, проведенными на эмбрионах рыб, в которых пионерные аксоны вырезались или удалялись (Kuwada 1986; Chitnis and Kuwada 1991; Chitnis et al. .1992 год; Пайк и др. 1992).
Покадровые исследования показали, что морфология конусов роста, поведение и динамика актина аксонов-первопроходцев отличаются от таковых у аксонов-последователей, которые менее сложны, растут с большей скоростью через точки выбора и имеют более высокую динамику актина (Bak и Фрейзер, 2003; Кулкарни и др., 2007). Роль пионерных аксонов в направлении последующих (из одного ядра или другого происхождения) в первую очередь подтверждается экспериментами, проведенными на эмбрионах рыб, в которых пионерные аксоны вырезались или удалялись (Kuwada 1986; Chitnis and Kuwada 1991; Chitnis et al. .1992 год; Пайк и др. 1992).
Только в конце 1980-х годов было заново открыто, что у всех видов позвоночных первые пути аксонов развиваются в чрезвычайно консервативной и стереотипной пространственно-временной последовательности (4). На очень ранней стадии пионерные аксоны устанавливают паттерны роста, которые сохраняются позже в развитии, такие как формирование продольного роста по сравнению с окружным, притяжение или отталкивание от средней линии и ростральная или каудальная ориентация. У позвоночных, несмотря на некоторую видовую изменчивость (Easter et al.1993 год; Чедоталь и др. 1995 год; Хьорт и Ки, 2002 г.; Баррейро-Иглесиас и др. 2008), сначала формируются два-три вентральных/базальных продольных тракта, которые тянутся от переднего и среднего мозга к заднему мозгу; к ним относятся тракт постоптической спайки (TPOC), медиальный продольный пучок (MLF) и нисходящий корешок мезэнцефального ядра тройничного нерва (MesV). Следующими формируются дорсо-вентральные тракты, аксоны которых позже присоединяются к продольным трактам или пересекают среднюю линию, образуя комиссуры.За последние 20 лет были предприняты большие усилия, чтобы понять механизмы, контролирующие развитие первых аксонов.
У позвоночных, несмотря на некоторую видовую изменчивость (Easter et al.1993 год; Чедоталь и др. 1995 год; Хьорт и Ки, 2002 г.; Баррейро-Иглесиас и др. 2008), сначала формируются два-три вентральных/базальных продольных тракта, которые тянутся от переднего и среднего мозга к заднему мозгу; к ним относятся тракт постоптической спайки (TPOC), медиальный продольный пучок (MLF) и нисходящий корешок мезэнцефального ядра тройничного нерва (MesV). Следующими формируются дорсо-вентральные тракты, аксоны которых позже присоединяются к продольным трактам или пересекают среднюю линию, образуя комиссуры.За последние 20 лет были предприняты большие усилия, чтобы понять механизмы, контролирующие развитие первых аксонов.
Схематическое изображение раннего аксонального каркаса у мышей и Xenopus . Сокращения: MesV, нисходящий путь мезэнцефального ядра тройничного нерва; MLF, медиальный продольный пучок; TPOC, тракт постоптической спайки; ПК, задняя спайка; IV, блоковый нерв; ММТ, маммилоталамический тракт; SOT, супраоптический тракт; AC, передняя спайка; ДВДТ, дорзовентральный диэнцефальный тракт; ПОС, постоптическая спайка; mes, мезэнцефалон; умирают, промежуточный мозг; тел, конечный мозг; ромб, ромбовидный мозг.
Классически нервная трубка эмбриона подразделяется в продольном направлении на три основных пузырька: прозэнцефалон, мезэнцефалон и ромбовидный мозг, каждый из которых разделен поперечными перетяжками. По мере развития в прозэнцефалоне добавляются новые поперечные сегменты, называемые просомерами, а в ромбэнцефалоне добавляются ромбомеры (Lumsden and Keynes 1989; Puelles and Rubenstein 2003; Kiecker and Lumsden 2005). Разграничение нейроэпителиальных доменов основывается на морфологических и молекулярных критериях, поскольку каждый домен экспрессирует четкую комбинацию факторов транскрипции и молекул клеточной адгезии, созданную специфическими морфогенами, такими как Fgfs, Shh и Wnts, которые обогащены на их границах.Интересно, что эти домены формируются до появления первых аксонов (HH9 у кур и E8.5 у мышей) (Sechrist and Bronner-Fraser 1991; Easter et al. 1993). У рыб, птиц и грызунов наблюдается поразительная, хотя и не абсолютная (Hjorth, Key, 2001), корреляция между участками роста пионерных аксонов и границами нейроэпителиальных доменов (Krauss et al. , 1991; Figdor, Stern, 1993; Макдональд и др., 1994). Следовательно, в заднем мозге сегментированный паттерн проекций моторных нейронов диктуется их ромбомерным происхождением () (Lumsden and Keynes 1989; Kiecker and Lumsden 2005).
, 1991; Figdor, Stern, 1993; Макдональд и др., 1994). Следовательно, в заднем мозге сегментированный паттерн проекций моторных нейронов диктуется их ромбомерным происхождением () (Lumsden and Keynes 1989; Kiecker and Lumsden 2005).
Плоское изображение ромбовидного мозга курицы Hh31, показывающее его раннюю сегментацию на 8 ромбомеров (r1-r8). Hox-код, характерный для каждого ромбомера или пары ромбомеров с нечетным и четным номером, обозначен цветовым кодом. Также представлены различные черепные моторные ядра и нервные корешки. Сокращения: IV — блоковое ядро; V, ядро тройничного нерва; VI, отводящее ядро; VII, лицевое ядро; IX — языкоглоточное ядро; X, ядро блуждающего нерва; XII, подъязычное ядро; Граница среднего и заднего мозга MHB; Mes, мезэнцефалон; ФП, напольная плита.Адаптировано из Kiecker and Lumsden, 2005.
Это выравнивание предполагает, что первые аксоны распознают сигналы управления, распределенные регионарным образом в нейроэпителии. В подтверждение этой идеи траектория нескольких ранних аксональных путей нарушена у мутантов рыб и мышей, лишенных факторов транскрипции Pax6 или гомолога Pax2 Noi (No-istmus, pax2. 1; Macdonald et al., 1994; Mastick et al. 1997; Уилсон и др., 1997). Аналогичным образом, у рыбок данио, лишенных cyclops (фактор, связанный с узлами), изменяется характер экспрессии нескольких транскрипционных факторов, что сопровождается дезорганизацией ранних аксональных путей (Macdonald et al.1994). Поскольку некоторыми нижестоящими мишенями этих транскрипционных факторов являются молекулы клеточной адгезии, такие как кадгерины и другие молекулы, направляющие аксоны (Nguyen Ba-Charvet et al., 1998; Andrews and Mastick, 2003; Geisen et al., 2008), различные свойства клеточной поверхности могут объяснять избирательное предпочтение пионерных аксонов для одних доменов или их исключение из других.
1; Macdonald et al., 1994; Mastick et al. 1997; Уилсон и др., 1997). Аналогичным образом, у рыбок данио, лишенных cyclops (фактор, связанный с узлами), изменяется характер экспрессии нескольких транскрипционных факторов, что сопровождается дезорганизацией ранних аксональных путей (Macdonald et al.1994). Поскольку некоторыми нижестоящими мишенями этих транскрипционных факторов являются молекулы клеточной адгезии, такие как кадгерины и другие молекулы, направляющие аксоны (Nguyen Ba-Charvet et al., 1998; Andrews and Mastick, 2003; Geisen et al., 2008), различные свойства клеточной поверхности могут объяснять избирательное предпочтение пионерных аксонов для одних доменов или их исключение из других.
В переднем мозге первые популяции аксонов пересекают границы, а не формируются вместе с ними. Например, переходный и гетерогенный слой нейронов, населяющих подпластину неокортекса (Ayoub and Kostovic 2009), является пионером кортико-таламической и таламокортикальной траектории, формирующей внутреннюю капсулу. Разрушение субпластины вызывает дефекты таламокортикального нацеливания (McConnell et al., 1989; Ghosh et al., 1990; Ghosh and Shatz, 1992). Временные контакты, образующиеся между нейронами субпластины и таламокортикальными аксонами, также необходимы для их правильной сегрегации и синаптического уточнения (Kanold et al. 2003). Первые аксоны также образуют первые проекции мозолистого тела и происходят из поясной коры, а не из неокортекса (Koester and O’Leary 1994; Rash and Richards 2001).Недавние доказательства их участия в формировании мозолистого тела указывают на участие в этом процессе передачи сигналов Npn1/Sema (Piper et al. 2009).
Разрушение субпластины вызывает дефекты таламокортикального нацеливания (McConnell et al., 1989; Ghosh et al., 1990; Ghosh and Shatz, 1992). Временные контакты, образующиеся между нейронами субпластины и таламокортикальными аксонами, также необходимы для их правильной сегрегации и синаптического уточнения (Kanold et al. 2003). Первые аксоны также образуют первые проекции мозолистого тела и происходят из поясной коры, а не из неокортекса (Koester and O’Leary 1994; Rash and Richards 2001).Недавние доказательства их участия в формировании мозолистого тела указывают на участие в этом процессе передачи сигналов Npn1/Sema (Piper et al. 2009).
В нескольких исследованиях предполагается, что гомеобоксные транскрипционные факторы сами по себе могут действовать как секретируемые факторы ближнего действия для пионерных аксонов, при этом новые данные демонстрируют поглощение и ретроградный транспорт транскрипционных факторов в соседние клетки (рассмотрено в Prochiantz and Joliot 2003 и Brunet et al. , 2007). Ростро-каудальное распределение аксонов сетчатки в среднем мозге соответствует градиенту транскрипционного фактора гомеобокса Engrailed, установленному Fgf8 (Itasaki and Nakamura 1996; Chen et al.2009), распространяющийся от границы среднего/заднего мозга. Активность Engrailed может включать нижестоящие мишени, лиганды ephrinA, но недавно было показано, что Engrailed может секретироваться тектальными клетками и интернализоваться аксонами сетчатки, индуцируя их поворот (Brunet et al. 2005). Сходным образом, некоторые тракты простираются вдоль доменов, экспрессирующих высокий уровень Otx2 (Nguyen Ba-Charvet et al. 1998), фактора транскрипции, который также может действовать вне клетки автономно во время развития мозга (Sugiyama et al. 2008). Непосредственное тестирование неклеточно-автономной роли гомеобокс-содержащих факторов транскрипции в раннем развитии аксонального тракта потребует блокирования их секреции или интернализации in vivo.
, 2007). Ростро-каудальное распределение аксонов сетчатки в среднем мозге соответствует градиенту транскрипционного фактора гомеобокса Engrailed, установленному Fgf8 (Itasaki and Nakamura 1996; Chen et al.2009), распространяющийся от границы среднего/заднего мозга. Активность Engrailed может включать нижестоящие мишени, лиганды ephrinA, но недавно было показано, что Engrailed может секретироваться тектальными клетками и интернализоваться аксонами сетчатки, индуцируя их поворот (Brunet et al. 2005). Сходным образом, некоторые тракты простираются вдоль доменов, экспрессирующих высокий уровень Otx2 (Nguyen Ba-Charvet et al. 1998), фактора транскрипции, который также может действовать вне клетки автономно во время развития мозга (Sugiyama et al. 2008). Непосредственное тестирование неклеточно-автономной роли гомеобокс-содержащих факторов транскрипции в раннем развитии аксонального тракта потребует блокирования их секреции или интернализации in vivo.
Другие недавние исследования показали, что первые аксоны в значительной степени реагируют на тот же набор направляющих молекул, что и более поздние аксоны. У рыбок данио Sema3D первоначально отталкивает аксоны MLF от переднего мозга и притягивает аксоны передней комиссуры к средней линии; однако на более поздней стадии он способствует фасцикуляции этих трактов, влияя на уровень L1-CAM на клеточной поверхности (Wolman et al. 2004; Wolman et al. 2007). У кур отталкивание Sema3F/нейропилина-2 также играет роль в сдерживании роста блоковых двигательных аксонов кур на границе среднего и заднего мозга (Watanabe et al.2004). У кур и мышей каудально выступающие продольные аксоны MLF и MesV экспрессируют Robo-рецепторы и растут между доменами, экспрессирующими высокие уровни Slit-лигандов (Molle et al. 2004; Farmer et al. 2008; Kastenhuber et al. 2009). У анамниот передача сигналов Slit/Robo контролирует фасцикуляцию аксонов TPOC, которые также распространяются на богатую Netrin область в базальных отделах переднего мозга (Wilson and Key 2006; Devine and Key 2008).
У рыбок данио Sema3D первоначально отталкивает аксоны MLF от переднего мозга и притягивает аксоны передней комиссуры к средней линии; однако на более поздней стадии он способствует фасцикуляции этих трактов, влияя на уровень L1-CAM на клеточной поверхности (Wolman et al. 2004; Wolman et al. 2007). У кур отталкивание Sema3F/нейропилина-2 также играет роль в сдерживании роста блоковых двигательных аксонов кур на границе среднего и заднего мозга (Watanabe et al.2004). У кур и мышей каудально выступающие продольные аксоны MLF и MesV экспрессируют Robo-рецепторы и растут между доменами, экспрессирующими высокие уровни Slit-лигандов (Molle et al. 2004; Farmer et al. 2008; Kastenhuber et al. 2009). У анамниот передача сигналов Slit/Robo контролирует фасцикуляцию аксонов TPOC, которые также распространяются на богатую Netrin область в базальных отделах переднего мозга (Wilson and Key 2006; Devine and Key 2008).
Факторы, обеспечивающие ростро-каудальную направленность в мозге, до сих пор неизвестны, но некоторые морфогены, такие как Shh, Wnt и Fgf, могут вносить свой вклад. Fgf8 регулирует формирование паттерна пионерных аксонов в переднем мозге (Shanmugalingam et al. 2000) и привлекает блоковые аксоны вдоль границы среднего/заднего мозга (Irving et al. 2002).
Fgf8 регулирует формирование паттерна пионерных аксонов в переднем мозге (Shanmugalingam et al. 2000) и привлекает блоковые аксоны вдоль границы среднего/заднего мозга (Irving et al. 2002).
МЕХАНИЗМЫ НАПРАВЛЕНИЯ АКСОНОВ В МОЗГЕ
Невероятная сложность мозга млекопитающих, а также нацеливание и рост аксонов на больших расстояниях требуют уникальной стратегии, позволяющей проводить проводку мозга во время развития. Это достигается за счет использования промежуточных целей. Для осуществления навигации аксонов на больших расстояниях система разбивается на более мелкие, более управляемые решения с помощью промежуточных мишеней, состоящих из глиальных клеток или промежуточных клеток-ориентиров.В заднем и среднем мозге это происходит через пластинку дна, временную глиоподобную структуру на вентральной срединной линии мозга. Однако между передним и задним мозгом существуют важные различия. В то время как в переднем мозге спайки ограничены ограниченным числом мест, комиссуральные аксоны широко распространены по всему заднему мозгу и спинному мозгу и имеют тенденцию к слиянию/фасцикуляции в четко определенные тракты только после того, как они пересекают срединную линию и принимают продольный способ роста. Это важное различие, вероятно, связано с наличием клеток пластинки дна, которые простираются от каудального конца спинного мозга до гипоталамуса и играют главную роль в формировании паттерна аксональных связей на этом уровне ЦНС посредством секреции хемоаттрактантов и хеморепеллентов. Напротив, комиссуральные аксоны в переднем мозге, по-видимому, проходят через очень специфические места.
Это важное различие, вероятно, связано с наличием клеток пластинки дна, которые простираются от каудального конца спинного мозга до гипоталамуса и играют главную роль в формировании паттерна аксональных связей на этом уровне ЦНС посредством секреции хемоаттрактантов и хеморепеллентов. Напротив, комиссуральные аксоны в переднем мозге, по-видимому, проходят через очень специфические места.
Комиссуральные и продольные проекции переднего мозга. И глиальные, и нейрональные структуры связаны с аксональными путями в головном мозге.( A – C ) изображают комиссуральные тракты на схемах горизонтальных сечений от спинного к брюшному. ( A и B ) представляют собой схемы головного мозга, тогда как C представляет собой вид головы снизу. С мозолистым телом (синий тракт на А) связаны глиальный клин, глия indusium griseum и пращевые клетки. Глия также связана со спайкой гиппокампа (фиолетовый тракт на А), передней спайкой (зеленый тракт на В) и перекрестом зрительных нервов (красные перекрестные волокна на С). В ( D ) показаны продольные пути, включая кортико-таламические, таламокортикальные, кортико-колликулярные и корково-спинномозговые пути, которые проходят через внутреннюю капсулу. С внутренней капсулой связаны коридорные клетки. Боковой обонятельный тракт (LOT) также показан на D вместе с клетками LOT. На всех схемах показаны срезы мозга или головы мыши на 18-й день эмбрионального развития.
В ( D ) показаны продольные пути, включая кортико-таламические, таламокортикальные, кортико-колликулярные и корково-спинномозговые пути, которые проходят через внутреннюю капсулу. С внутренней капсулой связаны коридорные клетки. Боковой обонятельный тракт (LOT) также показан на D вместе с клетками LOT. На всех схемах показаны срезы мозга или головы мыши на 18-й день эмбрионального развития.
В переднем мозге отсутствует структура пластинки дна, но присутствуют дополнительные, переходные срединные глиальные популяции, которые секретируют молекулы, сходные с пластинкой дна, в более каудальных областях нервной системы. .Срединные глиальные популяции связаны с каждым спаечным и перекрестным выступом в головном мозге (Silver et al. 1993). Эти глиальные популяции известны как «палисад» (перекрест зрительных нервов) (Marcus et al., 1995), «туннели» (передняя спайка и свод) (Pires-Neto et al., 1998; Braga-de-Souza and Lent, 2004; Lent et al. 2005), «клин» и «indusium griseum glia» (мозолистое тело) (Shu and Richards 2001; Shu et al. 2003b) (). Молекулы, экспрессируемые этими глиальными популяциями, включают Slits (Erskine et al.2000 г.; Пламп и др. 2002 г.; Шу и др. 2003c), Wnts (Keeble and Cooper 2006), Ephrins (Mendes et al. 2006; Williams et al., 2003), Draxin (Islam et al. 2009) и хондроитинсульфатные протеогликаны (Braga-de-Souza and Lent 2004) . Развитие этих глиальных популяций регулируется транскрипционными факторами, такими как гены Nfi (Shu et al. 2003a; Steele-Perkins et al. 2005; Barry et al. 2008) и передачей сигналов фактором роста фибробластов (Smith et al. 2006). Как и пластинка дна, каждая из этих популяций является временной и присутствует только во время развития путей аксонов, с которыми они связаны.
2003b) (). Молекулы, экспрессируемые этими глиальными популяциями, включают Slits (Erskine et al.2000 г.; Пламп и др. 2002 г.; Шу и др. 2003c), Wnts (Keeble and Cooper 2006), Ephrins (Mendes et al. 2006; Williams et al., 2003), Draxin (Islam et al. 2009) и хондроитинсульфатные протеогликаны (Braga-de-Souza and Lent 2004) . Развитие этих глиальных популяций регулируется транскрипционными факторами, такими как гены Nfi (Shu et al. 2003a; Steele-Perkins et al. 2005; Barry et al. 2008) и передачей сигналов фактором роста фибробластов (Smith et al. 2006). Как и пластинка дна, каждая из этих популяций является временной и присутствует только во время развития путей аксонов, с которыми они связаны.
В дополнение к транзиторным глиальным популяциям в головном мозге также был идентифицирован ряд транзиторных популяций нейронов, которые действуют как «клетки ориентира» или «клетки коридора» для аксонов. В развивающейся обонятельной системе основным эфферентным выступом обонятельной луковицы является латеральный обонятельный тракт (LOT), который содержит направляющие клетки, известные как клетки LOT (Tomioka et al. 2000; ). Клетки LOT мигрируют тангенциально и вентрально из неокортекса, чтобы остаться в латеральной части переднего мозга, где позже сформируется LOT.Миграция этих клеток управляется Netrin и Sema3F (Kawasaki et al. 2006; Ito et al. 2008), и развитие аксонов LOT зависит от присутствия этих клеток. Щели, секретируемые перегородкой, и несколько секретируемых семафоринов также играют ключевую роль в позиционировании LOT (de Castro et al., 1999; Nguyen Ba-Charvet et al., 1999; Nguyen-Ba-Charvet et al., 2002; Fouquet et al. 2007). В конечном мозге несколько других мигрирующих популяций нейронов участвуют в ведении аксонов. Подмозолистые пращевые клетки (Silver et al.1982 год; Шу и др. 2003b) и нейроны мозолистого тела (Riederer et al., 2004; Niquille et al., 2009) направляют аксоны мозолистого тела, а «коридорные» клетки внутренней капсулы направляют таламокортикальные аксоны в кору за счет экспрессии нейрегулина (Lopez — Бендито и др. 2006) (). Сходным образом, рано рожденные нейроны зрительного перекреста необходимы для направления аксонов ганглиозных клеток сетчатки по средней линии (Marcus and Mason 1995; Sretavan et al.
2000; ). Клетки LOT мигрируют тангенциально и вентрально из неокортекса, чтобы остаться в латеральной части переднего мозга, где позже сформируется LOT.Миграция этих клеток управляется Netrin и Sema3F (Kawasaki et al. 2006; Ito et al. 2008), и развитие аксонов LOT зависит от присутствия этих клеток. Щели, секретируемые перегородкой, и несколько секретируемых семафоринов также играют ключевую роль в позиционировании LOT (de Castro et al., 1999; Nguyen Ba-Charvet et al., 1999; Nguyen-Ba-Charvet et al., 2002; Fouquet et al. 2007). В конечном мозге несколько других мигрирующих популяций нейронов участвуют в ведении аксонов. Подмозолистые пращевые клетки (Silver et al.1982 год; Шу и др. 2003b) и нейроны мозолистого тела (Riederer et al., 2004; Niquille et al., 2009) направляют аксоны мозолистого тела, а «коридорные» клетки внутренней капсулы направляют таламокортикальные аксоны в кору за счет экспрессии нейрегулина (Lopez — Бендито и др. 2006) (). Сходным образом, рано рожденные нейроны зрительного перекреста необходимы для направления аксонов ганглиозных клеток сетчатки по средней линии (Marcus and Mason 1995; Sretavan et al. 1995).
1995).
Основная гипотеза в области управления аксонами заключалась в том, что конусы роста аксонов управляются молекулярными градиентами в развивающейся нервной системе.Хотя несколько анализов in vitro позволили проверить эту гипотезу (обзор Pujic et al. 2009), точные параметры, необходимые для аксонального хемотаксиса, все еще не установлены (Mortimer et al. 2009). Установлено, что по всей нервной оси действуют механизмы как притяжения, так и отталкивания, направляющие аксоны. В переднем, среднем и спинном мозге Draxin действует как репеллент (Islam et al., 2009; Naser et al., 2009), экспрессируемый крышей в спинном мозге и глиальным клином в переднем мозге.Молекулы семейств нейропилинов и семафоринов опосредуют направление как притяжения, так и отталкивания и играют важную роль в направлении и позиционировании мозолистого тела и передней спайки (Falk et al. 2005; Niquille et al. 2009; Piper et al. 2009; Хатанака и др., 2009). Netrin1 действует как аттрактант для кортикофугальных (Metin et al. 1997; Richards et al. 1997) и таламокортикальных путей (Braisted et al. 2000). Эти тракты, а также многие другие комиссуральные отростки в головном мозге поражаются как у мышей с мутацией Netrin1, так и у мышей с мутацией DCC (Serafini et al.1996 год; Фазели и др. 1997). В зрительной системе Netrin1 направляет аксоны диска зрительного нерва для входа в зрительный нерв (Deiner et al. 1997). Было показано, что разрезы действуют как хеморепульсивные сигналы для перекреста аксонов зрительного перекреста (Erskine et al. 2000; Plump et al. 2002), а также мозолистых аксонов (Shu and Richards 2001; Bagri et al. 2002; Shu et al. 2003c), но их роль в опосредовании управления другими комиссуральными проекциями переднего мозга тщательно не исследована. Было также показано, что в ряде систем Slits и их рецепторы, Robos, регулируют фасцикуляцию аксонных трактов.Как описано ранее, формирование пионерных аксонных трактов в головном мозге позволяет прибывающим позже аксонам использовать пионерные пути для направления путем фасцикуляции с этими аксонами.
1997; Richards et al. 1997) и таламокортикальных путей (Braisted et al. 2000). Эти тракты, а также многие другие комиссуральные отростки в головном мозге поражаются как у мышей с мутацией Netrin1, так и у мышей с мутацией DCC (Serafini et al.1996 год; Фазели и др. 1997). В зрительной системе Netrin1 направляет аксоны диска зрительного нерва для входа в зрительный нерв (Deiner et al. 1997). Было показано, что разрезы действуют как хеморепульсивные сигналы для перекреста аксонов зрительного перекреста (Erskine et al. 2000; Plump et al. 2002), а также мозолистых аксонов (Shu and Richards 2001; Bagri et al. 2002; Shu et al. 2003c), но их роль в опосредовании управления другими комиссуральными проекциями переднего мозга тщательно не исследована. Было также показано, что в ряде систем Slits и их рецепторы, Robos, регулируют фасцикуляцию аксонных трактов.Как описано ранее, формирование пионерных аксонных трактов в головном мозге позволяет прибывающим позже аксонам использовать пионерные пути для направления путем фасцикуляции с этими аксонами. Фасцикуляция происходит посредством аксон-аксонных взаимодействий и может быть опосредована молекулами клеточной адгезии (CAM), такими как NCAM, L1-CAM или TAG-1, или посредством гомофильных рецепторных взаимодействий между аксонами, опосредованными рецепторами Robo или Eph.
Фасцикуляция происходит посредством аксон-аксонных взаимодействий и может быть опосредована молекулами клеточной адгезии (CAM), такими как NCAM, L1-CAM или TAG-1, или посредством гомофильных рецепторных взаимодействий между аксонами, опосредованными рецепторами Robo или Eph.
Несмотря на очевидную неврологическую значимость, механизмы, контролирующие рост аксональных проекций нейронов с помощью биогенных аминовых нейротрансмиттеров, таких как катехоламины (норадреналин и дофамин), ацетилхолин или серотонин (5-гидрокситриптамин или 5-НТ), в значительной степени игнорировались.Недавние исследования онтогенеза этих систем у рыбок данио позволят использовать генетические методы для ответа на этот вопрос (McLean and Fetcho 2004; Kastenhuber et al. 2009; Lillesaar et al. 2009). У грызунов рост дофаминергических аксонов от среднего мозга к переднему мозгу, по-видимому, управляется классическими секретируемыми молекулами управления аксонами (см. обзор Van den Heuvel and Pasterkamp 2008). Дофаминергические нейроны экспрессируют рецепторы Robo и нейропилина (Nakamura et al., 2000; Marillat et al., 2000).2002 г.; Эрнандес-Монтьель и др. 2008) и реагируют на несколько белков Semaphorins и Slit. Ростральный рост дофаминергических аксонов находится под влиянием отталкивающего градиента Sema3F, регулируемого Fgf8, возникающего на границе среднего и заднего мозга (Nakamura et al., 2000; Kolk et al., 2009; Yamauchi et al., 2009). Эти аксоны также управляются привлекательной активностью Sema3A и Sema3C, продуцируемой в промежуточном мозге и стриатуме (Hernandez-Montiel et al. 2008; Yamauchi et al. 2009), и Sema 3F, действующим через Npn2 в медиальной префронтальной коре (Kolk et al. др., 2009). Дофаминергические аксоны также отвечают in vitro на эксплантаты пластинки дна и белки Netrin1 и Slit (Lin et al. 2005). Наконец, дофаминергические проекции, которые в первую очередь являются ипсилатеральными, дефасцикулируют и пересекают срединную линию у мышей с двойным нокаутом Slit1/Slit2 (Bagri et al.
Дофаминергические нейроны экспрессируют рецепторы Robo и нейропилина (Nakamura et al., 2000; Marillat et al., 2000).2002 г.; Эрнандес-Монтьель и др. 2008) и реагируют на несколько белков Semaphorins и Slit. Ростральный рост дофаминергических аксонов находится под влиянием отталкивающего градиента Sema3F, регулируемого Fgf8, возникающего на границе среднего и заднего мозга (Nakamura et al., 2000; Kolk et al., 2009; Yamauchi et al., 2009). Эти аксоны также управляются привлекательной активностью Sema3A и Sema3C, продуцируемой в промежуточном мозге и стриатуме (Hernandez-Montiel et al. 2008; Yamauchi et al. 2009), и Sema 3F, действующим через Npn2 в медиальной префронтальной коре (Kolk et al. др., 2009). Дофаминергические аксоны также отвечают in vitro на эксплантаты пластинки дна и белки Netrin1 и Slit (Lin et al. 2005). Наконец, дофаминергические проекции, которые в первую очередь являются ипсилатеральными, дефасцикулируют и пересекают срединную линию у мышей с двойным нокаутом Slit1/Slit2 (Bagri et al. 2002).
2002).
Развитие дофаминергических проекций у эмбриона мыши. Дофаминергические аксоны берут начало из черной субстанции (SN) и вентральной области покрышки (VTA) в среднем мозге и иннервируют стриатум и кору.( A ) и ( B ) показывают развитие этого тракта на E11 и P0 соответственно. Они растут рострально под действием отталкивающего Sema3F, секретируемого на границе среднего и заднего мозга (MHB, C; C представляет собой увеличение области, заключенной в рамку в [ A ]). Градиент Sema3F контролируется Fgf8. Секретируемые репелленты из среднего и промежуточного мозга/таламуса ( B , C ) поддерживают дофаминергические аксоны вентрально, тогда как факторы, секретируемые из полосатого тела, привлекают их.( D ) Дофаминергические аксоны в основном выступают ипсилатерально и удерживаются вдали от средней линии с помощью Slits и других репеллентов. Сокращения: os — зрительный стебель; тел, конечный мозг; умирают, промежуточный мозг; mes, мезэнцефалон; ромб, ромбовидный мозг; Стри, полосатое тело; Таламус, таламус. Модифицировано из Yamauchi et al., 2009 и Van den Heuvel and Pasterkamp, 2008.
Модифицировано из Yamauchi et al., 2009 и Van den Heuvel and Pasterkamp, 2008.
Проекции аксонов обычно формируются стереотипно. Однако недавно в исследовании на мутантных мышах Sema6A была отмечена замечательная пластичность мозга (Little et al.2009). Поскольку большинство моделей мышей с мутациями в генах управления аксонами умирают при рождении, было мало возможностей исследовать, что происходит с неправильно нацеленными аксонами у взрослых животных. Однако у мутанта Sema6A , который доживает до зрелого возраста, некоторые таламические аксоны из латерального коленчатого ядра эктопически проецируются в неокортекс через верхние слои. Несмотря на то, что первичная зрительная зона уменьшена, эктопические проекции остаются у взрослых (Little et al.2009). Это открытие иллюстрирует пластичность нацеливания аксонов и способность сохранять функциональные эктопические проекции во взрослом возрасте.
КОНТРАЛАТЕРАЛЬНЫЕ И ИПСИЛАТЕРАЛЬНЫЕ ПУТИ В МОЗГЕ
Мозг млекопитающих имеет проводную структуру, основанную на функциональной специфичности, при этом различные области мозга, отвечающие за одну и ту же функциональную модальность, соединены между собой. Установление этой функциональной специфичности контролируется экспрессией генов на ранних стадиях развития, за которой следуют более поздние механизмы уточнения, зависящие от активности.
Установление этой функциональной специфичности контролируется экспрессией генов на ранних стадиях развития, за которой следуют более поздние механизмы уточнения, зависящие от активности.
В переднем мозге недавние исследования показывают, что транскрипционный контроль специфичности аксонов происходит, когда нейроны рождаются в вентрикулярной и субвентрикулярной зонах (VZ и SVZ, соответственно) неокортекса. Неокортикальная связь включает в себя как проекционные нейроны VZ и SVZ, так и интернейроны, рождающиеся в вентральной части переднего мозга и корковом крае (rev. Pierani and Wassef 2009). Проекционные нейроны связывают мозг на большие расстояния и обеспечивают связь между различными областями мозга, специализирующимися на одной и той же сенсомоторной модальности.Как проекционные нейроны находят свои цели в мозгу — важный вопрос нейробиологии. Недавнее понимание пришло из исследований, идентифицирующих факторы транскрипции, которые придают специфичность слоев и соединений проекционным нейронам в коре (rev. Molyneaux et al. 2007 и Leone et al. 2008). Некоторые из наиболее важных экспериментов в этом отношении связаны с манипулированием экспрессией генов, что приводит к переспецификации аксонов в другой тип проекционных нейронов. Например, Ctip2, Fezf2 и Sox5 определяют субцеребральные проекции, а Satb2 определяет мозолистые проекции в этой системе (Molyneaux et al.2005 г.; Алкамо и др. 2008 г.; Арлотта и др. 2008 г.; Британова и др. 2008 г.; Чен и др. 2008 г.; Кван и др. 2008 г.; Лай и др. 2008). В дополнение к этим исследованиям факторы транскрипции также регулируют региональные различия в человеческом мозге (Johnson et al., 2009), а также специфичность пути аксонов в других трактах, включая ретино-тектальную систему и таламокортикальную проекцию, а также моторику спинного мозга. — и проекции сенсорных нейронов (рассмотрено в Polleux et al. 2007). Сходным образом, в заднем и спинном мозге факторы транскрипции, такие как белки, содержащие гомеодомен, действуют выше многих рецепторов и лигандов наведения аксонов (см.
Molyneaux et al. 2007 и Leone et al. 2008). Некоторые из наиболее важных экспериментов в этом отношении связаны с манипулированием экспрессией генов, что приводит к переспецификации аксонов в другой тип проекционных нейронов. Например, Ctip2, Fezf2 и Sox5 определяют субцеребральные проекции, а Satb2 определяет мозолистые проекции в этой системе (Molyneaux et al.2005 г.; Алкамо и др. 2008 г.; Арлотта и др. 2008 г.; Британова и др. 2008 г.; Чен и др. 2008 г.; Кван и др. 2008 г.; Лай и др. 2008). В дополнение к этим исследованиям факторы транскрипции также регулируют региональные различия в человеческом мозге (Johnson et al., 2009), а также специфичность пути аксонов в других трактах, включая ретино-тектальную систему и таламокортикальную проекцию, а также моторику спинного мозга. — и проекции сенсорных нейронов (рассмотрено в Polleux et al. 2007). Сходным образом, в заднем и спинном мозге факторы транскрипции, такие как белки, содержащие гомеодомен, действуют выше многих рецепторов и лигандов наведения аксонов (см. обзор Chédotal and Rijli 2009).
обзор Chédotal and Rijli 2009).
В дополнение к спецификации слоев и типов клеток решающую роль в определении развития путей аксонов играют направляющие молекулы аксонов в головном мозге. Формирование как ипсилатеральных, так и контралатеральных аксонных путей позволяет мозгу интегрировать сенсорную и моторную информацию из окружающей среды с обеих сторон тела и выполнять соответствующие поведенческие реакции. Точно так же эволюционно консервативные молекулы, такие как Slits, IgCAMs, Netrins, Semaphorins и Ephrins, регулируют образование путей аксонов в мозге млекопитающих.Имеются примеры молекул из каждого семейства наведения, которые регулируют образование как ипсилатеральных, так и контралатеральных проекций: таким образом, начальное направление роста может определяться не молекулами наведения аксонов, а скорее регуляцией транскрипции. Эти факторы транскрипции могут регулировать экспрессию рецепторов в определенные периоды развития, позволяя аксонам управлять внешними сигналами (например, в зрительной системе) (Petros et al. 2008), но пока неясно, как осуществляется регуляция транскрипции. способны придавать тракту аксона и специфичность направления в большинстве систем мозга (Chédotal and Rijli 2009).
2008), но пока неясно, как осуществляется регуляция транскрипции. способны придавать тракту аксона и специфичность направления в большинстве систем мозга (Chédotal and Rijli 2009).
КАК НЕЙРОНЫ РАСПОЛОЖЕНЫ И СВЯЗЫВАЮТСЯ СО СВОЕЙ МИШЕНЬЮ В МОЗГЕ МЛЕКОПИТАЮЩИХ
использовались для исследования роли механизмов, зависящих от активности, в ведении аксонов и уточнении формирования карты. Наше понимание как молекулярных, так и зависящих от активности механизмов основано на этих экспериментальных парадигмах.
Что касается направления аксонов, не все нейроны рождаются одинаковыми.Создание синапсов на нужных клетках-мишенях — проблема чрезвычайной сложности, зависящая от типа нейрона. С одной стороны, это не проблема для нейронов, не имеющих аксона, таких как большинство амакриновых клеток сетчатки и зернистых клеток обонятельной луковицы. С другой стороны, это особенно сложная задача для аксонов, формирующих соединения «точка-точка» с уникальной отдаленной клеткой-мишенью.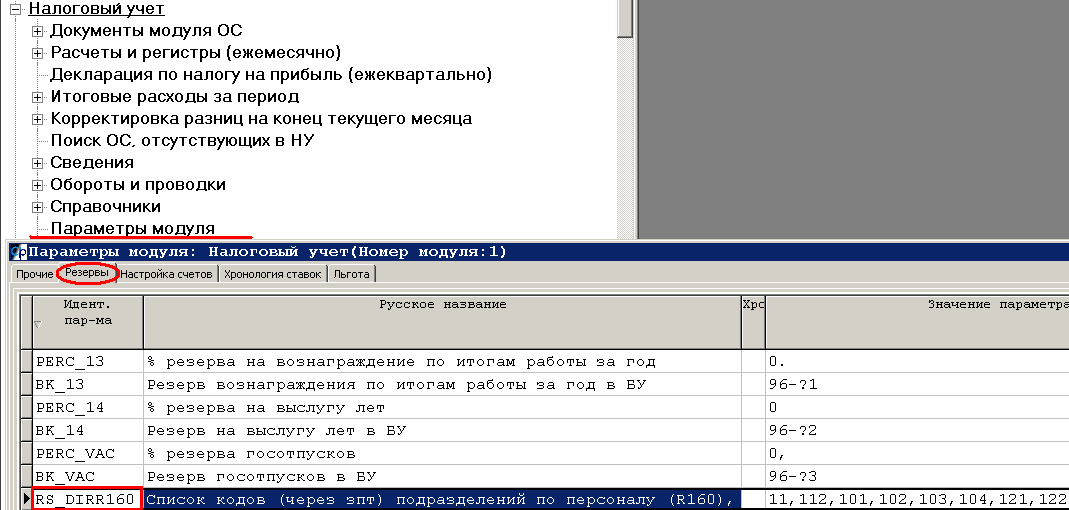 Более того, распределение большинства окончаний аксонов на их нейроне-мишени не является случайным, а ограничено специфическими субклеточными компартментами, такими как тело клетки, дендрит, шипы и аксон.Здесь мы суммируем различные молекулярные механизмы, которые контролируют окончательное нацеливание на некоторые классы нейронов.
Более того, распределение большинства окончаний аксонов на их нейроне-мишени не является случайным, а ограничено специфическими субклеточными компартментами, такими как тело клетки, дендрит, шипы и аксон.Здесь мы суммируем различные молекулярные механизмы, которые контролируют окончательное нацеливание на некоторые классы нейронов.
Прежде всего можно провести важное различие между интернейронами, которые контактируют с клетками-мишенями в непосредственной близости от них, и проекционными нейронами, цели которых могут находиться на расстоянии нескольких миллиметров. Аксоны от различных типов интернейронов не растут и образуют синапсы случайным образом, а разветвляются по определенным паттернам и слоям, что лучше всего видно на примере коры головного мозга (Huang et al. 2007; Ascoli et al.2008 г.; Батиста-Брито и Фишелл, 2009).
Недавние исследования показали, что молекулы клеточной адгезии суперсемейства иммуноглобулинов (IgCAM) направляют аксоны нескольких классов интернейронов в переднем и заднем мозге. В молекулярном слое мозжечка два типа ГАМКергических тормозных интернейронов, корзинчатые клетки и звездчатые клетки, иннервируют одну и ту же мишень, клетку Пуркинье, которая является единственным выходным нейроном коры мозжечка (Sotelo 2008). В то время как аксоны звездчатых клеток иннервируют только гладкую поверхность проксимальных дендритов клеток Пуркинье, корзинчатые клетки иннервируют сому клеток Пуркинье и аксон на уровне начального сегмента аксона, формируя характерные «пинсо» образования (Sotelo 2008).Два белка семейства L1-CAM IgCAM контролируют дифференцированное нацеливание на звездчатые и корзинчатые клетки (Ango et al. 2004; Ango et al. 2008). Было показано, что нейрофасцин 186 градиентно экспрессируется на теле клетки Пуркинье и увеличивается в начальном сегменте аксона, где он связывается с AnkyrinG (Ango et al. 2004). Аксоны корзинчатых клеток не могут должным образом нацеливаться на начальный сегмент аксона при устранении градиента NF186, например, у мышей с нокаутом AnkyrinG или после экспрессии доминантно-негативной формы нейрофасцина в клетках Пуркинье (14).
В молекулярном слое мозжечка два типа ГАМКергических тормозных интернейронов, корзинчатые клетки и звездчатые клетки, иннервируют одну и ту же мишень, клетку Пуркинье, которая является единственным выходным нейроном коры мозжечка (Sotelo 2008). В то время как аксоны звездчатых клеток иннервируют только гладкую поверхность проксимальных дендритов клеток Пуркинье, корзинчатые клетки иннервируют сому клеток Пуркинье и аксон на уровне начального сегмента аксона, формируя характерные «пинсо» образования (Sotelo 2008).Два белка семейства L1-CAM IgCAM контролируют дифференцированное нацеливание на звездчатые и корзинчатые клетки (Ango et al. 2004; Ango et al. 2008). Было показано, что нейрофасцин 186 градиентно экспрессируется на теле клетки Пуркинье и увеличивается в начальном сегменте аксона, где он связывается с AnkyrinG (Ango et al. 2004). Аксоны корзинчатых клеток не могут должным образом нацеливаться на начальный сегмент аксона при устранении градиента NF186, например, у мышей с нокаутом AnkyrinG или после экспрессии доминантно-негативной формы нейрофасцина в клетках Пуркинье (14). Гомофильные взаимодействия между отростками глии Бергмана и аксонами звездчатых клеток, по-видимому, направляют эти аксоны к дендритам клеток Пуркинье (Ango et al. 2008). Это включает в себя близкий гомолог L1 (CHL1), потому что у мышей с нокаутом CHL1 аксоны звездчатых клеток не могут должным образом иннервировать дендриты клеток Пуркинье. В сетчатке гомофильные взаимодействия с участием DsCAMs и Sidekick IgCAMs координируют точную проводку подмножеств биполярных нейронов, амакриновых клеток и ганглиозных клеток сетчатки во внутреннем плексиформном слое (Yamagata and Sanes 2008).
Гомофильные взаимодействия между отростками глии Бергмана и аксонами звездчатых клеток, по-видимому, направляют эти аксоны к дендритам клеток Пуркинье (Ango et al. 2008). Это включает в себя близкий гомолог L1 (CHL1), потому что у мышей с нокаутом CHL1 аксоны звездчатых клеток не могут должным образом иннервировать дендриты клеток Пуркинье. В сетчатке гомофильные взаимодействия с участием DsCAMs и Sidekick IgCAMs координируют точную проводку подмножеств биполярных нейронов, амакриновых клеток и ганглиозных клеток сетчатки во внутреннем плексиформном слое (Yamagata and Sanes 2008).
Развитие аксонов корзинчатых клеток. ( A ) оригинальный рисунок Сантьяго Рамона-и-Кахаля корзинчатой клетки мозжечка ( B ), помеченный окрашиванием по Гольджи, демонстрирующий характерные аксональные ветви (а), образования «пинсо» вокруг тела клетки Пуркинье и аксона. ( B ) Синапс аксонов корзинчатых клеток преимущественно находится на начальном сегменте аксона клеток Пуркинье (AIS) под влиянием градиента нейрофасцина 186, стабилизированного анкирином G. У мышей, нокаутированных по анкирину G, градиент нейрофасцина отменяется, и корзинчатая клетка аксоны не образуют синапсов преимущественно на AIS.Сокращения: ML — молекулярный слой; PCL, слой клеток Пуркинке; GCL, зернистый клеточный слой. А, рисунок Кахаля. Оригинал хранится в Instituto Cajal (CSIC), Мадрид (Испания). B взято из Huang et al., 2007.
У мышей, нокаутированных по анкирину G, градиент нейрофасцина отменяется, и корзинчатая клетка аксоны не образуют синапсов преимущественно на AIS.Сокращения: ML — молекулярный слой; PCL, слой клеток Пуркинке; GCL, зернистый клеточный слой. А, рисунок Кахаля. Оригинал хранится в Instituto Cajal (CSIC), Мадрид (Испания). B взято из Huang et al., 2007.
Проведение длинных проекционных аксонов также представляет собой довольно сложную задачу в зависимости от типа нейронов. Таламические аксоны направляются к определенным областям и слоям коры за счет экспрессии молекул, таких как эфрин/Eph (Uziel et al. 2005), но также сортируются во внутренней капсуле перед входом в кору посредством передачи сигналов эфрина/Eph и Netrin1. Дюфур и др.2003 г.; Пауэлл и др. 2008). Корково-спинномозговые аксоны, которые составляют самые длинные аксональные отростки в нервной системе, управляются экспрессией множественных направляющих молекул, включая экспрессию градиента Wnt, который направляет их назад (Liu et al. 2005; Canty and Murphy 2008).
2005; Canty and Murphy 2008).
Топография аминергических выступов достаточно рыхлая. Холинергические нейроны из базальных отделов переднего мозга (медиальная перегородка, диагональная полоса Брока, безымянная субстанция и бледный шар) интенсивно иннервируют кору головного мозга (Gould et al.1991). Дофаминергические нейроны среднего мозга из черной субстанции и вентральной области покрышки проецируются в первую очередь в стриатум и неокортекс. Плотность дофаминергических входов варьируется между слоями и областями коры, но четкой специфичности нет. Это также относится к норадренегическим проекциям из голубого пятна и серотонинергическим проекциям из ствола головного мозга, которые диффузно проецируются по всему головному и спинному мозгу (Gaspar et al. 2003).
В отличие от аминергических нейронов, большинство проекционных нейронов создают проекции с точно определенным паттерном при входе в домен-мишень.Хотя механизмы, зависящие от активности, контролируют окончательное уточнение этих проекций, их нацеливание на специфические слои и нейроны, а также их внутриклеточная локализация в первую очередь контролируются внешними сигналами.
В развивающемся гиппокампе аксоны энторинальной коры синапсируются на дистальной части дендритов гранулярных клеток в зубчатой извилине, тогда как проксимальная область дендрита нацелена на комиссуральные/ассоциативные аксоны из контралатерального гиппокампа (Super and Soriano 1994; Форстер и др.2006). До контакта с гранулярными клетками энторинальные аксоны также проецируются на временную популяцию нейронов, клетки Кахаля-Ретциуса. Предотвращение этого контакта сильно мешает их окончательному нацеливанию (Del Rio et al. 1997) и может быть вовлечен Reelin, секретируемый клетками Cajal-Retzius.
Другие примеры, иллюстрирующие роль молекул наведения аксонов в формировании проекционной карты, представлены в O’Leary (2010). Интересно, что появляется все больше данных, свидетельствующих о том, что на развитие двухточечных проекций влияют аминергические системы, в частности серотонин.Серотонин экспрессируется на ранних стадиях эмбрионального развития, и растущие аксоны могут высвобождать его до синаптогенеза. Сегрегация таламокортикальных аксонов в бочкообразном поле соматосенсорной коры блокируется у нескольких линий генетически модифицированных мышей с повышенным уровнем серотонина (Gaspar et al. 2003) (). Это также влияет на сегрегацию ипсилатеральных/контралатеральных глазных входов в латеральном коленчатом теле. В более раннем возрасте серотонин также может влиять на направление таламокортикальных аксонов, переводя активность Netrin1 с притяжения на отталкивание (Bonnin et al.2007). Действие серотонина на развивающиеся аксоны, по-видимому, в значительной степени опосредовано G i/o -сопряженными 5-HT 1B рецепторами, которые вызывают снижение уровня цАМФ.
Сегрегация таламокортикальных аксонов в бочкообразном поле соматосенсорной коры блокируется у нескольких линий генетически модифицированных мышей с повышенным уровнем серотонина (Gaspar et al. 2003) (). Это также влияет на сегрегацию ипсилатеральных/контралатеральных глазных входов в латеральном коленчатом теле. В более раннем возрасте серотонин также может влиять на направление таламокортикальных аксонов, переводя активность Netrin1 с притяжения на отталкивание (Bonnin et al.2007). Действие серотонина на развивающиеся аксоны, по-видимому, в значительной степени опосредовано G i/o -сопряженными 5-HT 1B рецепторами, которые вызывают снижение уровня цАМФ.
Серотонин влияет на разветвление аксонов во время развития. В слое IV соматосенсорной коры таламические аксоны передают сенсорную информацию от одного и того же кластера усов и разветвляются в одном домене, называемом «стволом». Когда уровень серотонина увеличивается во время развития, как это происходит у мышей с нокаутом MaoA и SERT, бочкообразное поле не формируется и окончания таламических аксонов, соответствующие отдельным усам, перекрываются. Адаптировано из Gaspar et al., 2003.
Адаптировано из Gaspar et al., 2003.
Аналогичная роль других биогенных аминов еще не продемонстрирована, но ацетилхолин, продуцируемый некоторыми амакриновыми клетками, необходим для создания визуальных проекционных карт (Cang et al. 2008; Huberman et al. 2008 г.). Ацетилхолин привлекает аксоны спинного мозга в тесте поворота Xenopus (Zheng et al. 1994), что позволяет предположить, что этот нейротрансмиттер может влиять на направление аксонов во многих областях мозга. Интересно, что другие циркулирующие белки, такие как эндоканнабиноиды (Berghuis et al.2007) и лептин (Bouret et al. 2004) также могут влиять на направление аксонов.
ТЕКУЩИЕ НАПРАВЛЕНИЯ В ПРОВОДКЕ МОЗГА МЛЕКОПИТАЮЩИХ
Описанная здесь работа продемонстрировала некоторые всеобъемлющие механистические принципы в проводке мозга млекопитающих. Первые аксоны впервые подготовили почву для общей аксональной карты мозга. Во-вторых, популяции нейронов генерируются и специфицируются экспрессией факторов транскрипции, при этом эти нейроны посылают аксоны, которые растут к своим конечным мишеням за счет использования промежуточных мишеней, таких как глиальные и коридорные клетки, вдоль пути. Специфичность мишени и синапса в области конечной мишени координируется как посредством молекулярных, так и зависящих от активности механизмов. Эти последовательные механизмы, работающие вместе в нейронах, формирующих разные, но жизненно важные функции мозга, лежат в основе функциональной схемы мозга взрослого человека. Будущие направления в этой области будут включать впечатляющие технические достижения в наблюдении одиночных нейронов и нейронных цепей с помощью флуоресцентных меток или с помощью технологий визуализации мозга (например, см. линии, доступные от консорциума Gensat) (Gong et al.2003 г. и технология Brainbow; Ливет и др. 2007) (). В настоящее время также предпринимаются скоординированные усилия по картированию связей нейронов в мозге многих организмов (Bohland et al. 2009), особенно мыши и макаки, а также по разработке инструментов для картирования связей нейронов человека в беспрецедентном масштабе. Это исследование приведет к еще одному скачку в нашем понимании структуры и функций мозга, поскольку мы сможем анализировать нейронные цепи как на микроскопическом, так и на системном уровнях способами, которые ранее были невозможны.
Специфичность мишени и синапса в области конечной мишени координируется как посредством молекулярных, так и зависящих от активности механизмов. Эти последовательные механизмы, работающие вместе в нейронах, формирующих разные, но жизненно важные функции мозга, лежат в основе функциональной схемы мозга взрослого человека. Будущие направления в этой области будут включать впечатляющие технические достижения в наблюдении одиночных нейронов и нейронных цепей с помощью флуоресцентных меток или с помощью технологий визуализации мозга (например, см. линии, доступные от консорциума Gensat) (Gong et al.2003 г. и технология Brainbow; Ливет и др. 2007) (). В настоящее время также предпринимаются скоординированные усилия по картированию связей нейронов в мозге многих организмов (Bohland et al. 2009), особенно мыши и макаки, а также по разработке инструментов для картирования связей нейронов человека в беспрецедентном масштабе. Это исследование приведет к еще одному скачку в нашем понимании структуры и функций мозга, поскольку мы сможем анализировать нейронные цепи как на микроскопическом, так и на системном уровнях способами, которые ранее были невозможны.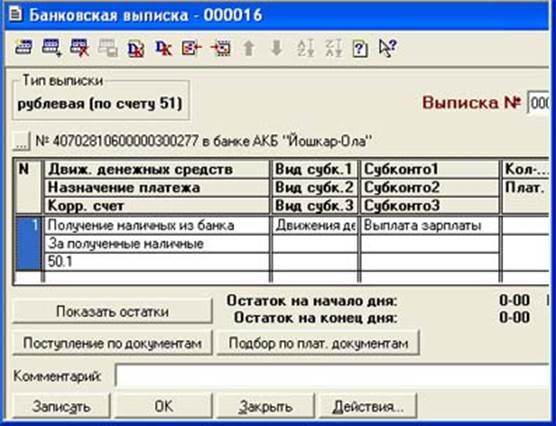
Достижения в методах маркировки путей и цепей аксонов. ( A ) Маркировка карбоцианиновым красителем в развивающемся мозге. Мозолистые аксоны, меченные DiI, показаны красным (стрелка на А). Мечение проводили на фиксированном эмбриональном мозге мыши 17-го дня. (изображение предоставлено доктором Селин Плачес, Мэрилендский университет). ( B ) Маркировка нейронов в мозгу «мозговой лук» мыши. Различные нейроны визуализируются с разными оттенками цвета, генерируемыми рекомбинацией Cre/loxP у трансгенных мышей (изображение предоставлено доктором Джеффом Лихтманом и доктором Тамили Вайсман, Гарвардский университет).( C ) Диффузионно-взвешенное (30 направлений) магнитно-резонансное изображение, полученное при 16,4 Тесла — цветовая карта, демонстрирующая комиссуральные пути в средней сагиттальной проекции. Основываясь на своей ориентации, комиссуральные волокна были отмечены красным цветом, включая мозолистое тело (стрелка на С) и переднюю спайку (стрелка на С). ( D и E ) представляют собой трактографические изображения высокого углового разрешения (HARDI/q-ball). В D области интереса (ROI) были выбраны по всему мозгу, с показанными аксонными путями, которые проходят через среднюю линию.E демонстрирует более избирательное размещение областей интереса: одна на средней линии в пределах передней комиссуры (стрелки на E изображают как переднее, так и заднее плечо передней комиссуры, которые проходят через область интереса по средней линии), а одна в заднем мозге на уровне срединная линия в пределах средней ножки мозжечка и поперечных волокон моста (стрелка на E). Изображения в C-E предоставлены доктором Ниоманом Курниаваном и доктором Рэндалом Молдричем (Университет Квинсленда). Масштабная линейка в E = 400 мкм в A, 80 мкм в B, 2 мм в C и E и 1.35 мм в D.
( D и E ) представляют собой трактографические изображения высокого углового разрешения (HARDI/q-ball). В D области интереса (ROI) были выбраны по всему мозгу, с показанными аксонными путями, которые проходят через среднюю линию.E демонстрирует более избирательное размещение областей интереса: одна на средней линии в пределах передней комиссуры (стрелки на E изображают как переднее, так и заднее плечо передней комиссуры, которые проходят через область интереса по средней линии), а одна в заднем мозге на уровне срединная линия в пределах средней ножки мозжечка и поперечных волокон моста (стрелка на E). Изображения в C-E предоставлены доктором Ниоманом Курниаваном и доктором Рэндалом Молдричем (Университет Квинсленда). Масштабная линейка в E = 400 мкм в A, 80 мкм в B, 2 мм в C и E и 1.35 мм в D.
Визуализация всего мозга для понимания вопросов системного уровня также развивается быстрыми темпами. Магнитно-резонансная томография использовалась для изучения общей анатомии живого человеческого мозга.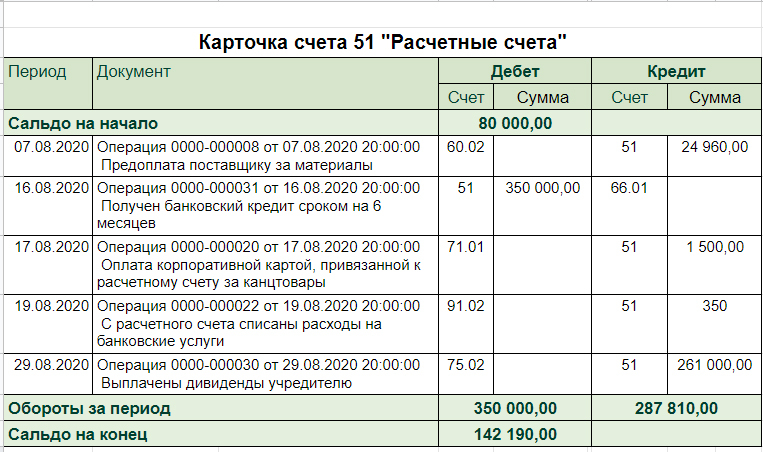 Недавние достижения в этой области привели к развитию неинвазивной аксональной трактографии и отслеживания путей. Тензор диффузии и взвешенная по диффузии визуализация позволяют кодировать аксональные тракты в головном мозге в зависимости от ориентации волокон (Behrens et al. 2003) (). Новые методы, известные как диффузионная визуализация с высоким угловым разрешением (HARDI) и трактография q-Ball, обеспечивают еще большую точность, особенно в областях пересечения волокон.Подобные методы можно использовать как на фиксированных тканях, так и на живом мозге, и они могут обеспечить невероятную трехмерную реконструкцию аксональных путей и их связи с другими аксональными путями в головном мозге. Недавнее использование этой техники у пациентов с мозолистым гипогенезом продемонстрировало значительный набор различий в проводке мозга между пациентами со схожими макроанатомическими особенностями (Wahl et al., 2009) и даже выявило образование эктопических связей у этих пациентов (Tovar- Молл и др.
Недавние достижения в этой области привели к развитию неинвазивной аксональной трактографии и отслеживания путей. Тензор диффузии и взвешенная по диффузии визуализация позволяют кодировать аксональные тракты в головном мозге в зависимости от ориентации волокон (Behrens et al. 2003) (). Новые методы, известные как диффузионная визуализация с высоким угловым разрешением (HARDI) и трактография q-Ball, обеспечивают еще большую точность, особенно в областях пересечения волокон.Подобные методы можно использовать как на фиксированных тканях, так и на живом мозге, и они могут обеспечить невероятную трехмерную реконструкцию аксональных путей и их связи с другими аксональными путями в головном мозге. Недавнее использование этой техники у пациентов с мозолистым гипогенезом продемонстрировало значительный набор различий в проводке мозга между пациентами со схожими макроанатомическими особенностями (Wahl et al., 2009) и даже выявило образование эктопических связей у этих пациентов (Tovar- Молл и др.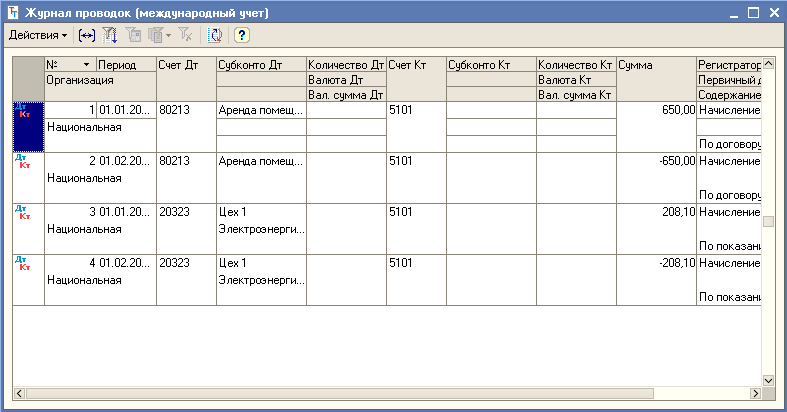 2007). Эта работа подготовила почву для беспрецедентного понимания и измерения пластичности человеческого мозга во время развития и после травмы или болезни, а также для возможности связать фактическую связь мозга с поведением человека.
2007). Эта работа подготовила почву для беспрецедентного понимания и измерения пластичности человеческого мозга во время развития и после травмы или болезни, а также для возможности связать фактическую связь мозга с поведением человека.
ПРИЗНАТЕЛЬНОСТЬ
Alain Chédotal поддерживается Национальным институтом здоровья и медицинских исследований, Национальным агентством по исследованиям (программы Blanc и MNP) и Фондом медицинских исследований (программа équipe FRM).Линда Ричардс поддерживается стипендией старшего научного сотрудника Национального совета по здравоохранению и медицинским исследованиям Австралии. Мы благодарим Джона Бейсдена и Яна Глиддена за помощь с графикой и Роуэна Твидейла за критическое прочтение текста.
Сноски
Редакторы: Marc Tessier-Lavigne и Alex L. Kolodkin
Дополнительные перспективы нейронной навигации доступны на сайте www.cshperspectives.org , Гроссшедл Р., МакКоннелл С.К. 2008.Satb2 регулирует идентичность каллозальных проекционных нейронов в развивающейся коре головного мозга. Neuron 57:364–377 [PubMed] [Google Scholar]
Neuron 57:364–377 [PubMed] [Google Scholar]
 J Neurosci 28:622–632 [бесплатная статья PMC] [PubMed] [Google Scholar]
J Neurosci 28:622–632 [бесплатная статья PMC] [PubMed] [Google Scholar] Development 130:4999–5008 [PubMed] [Google Scholar]
Development 130:4999–5008 [PubMed] [Google Scholar] Nat Neurosci 6:750–757 [PubMed] [Google Scholar]
Nat Neurosci 6:750–757 [PubMed] [Google Scholar] Временная и пространственная регуляция хондроитинсульфата, радиальных глиальных клеток, растущих комиссуральных аксонов и других эфферентов гиппокампа у развивающихся хомячков.J Comp Neurol 468:217–232 [PubMed] [Google Scholar]
Временная и пространственная регуляция хондроитинсульфата, радиальных глиальных клеток, растущих комиссуральных аксонов и других эфферентов гиппокампа у развивающихся хомячков.J Comp Neurol 468:217–232 [PubMed] [Google Scholar] Фактор транскрипции Engrailed-2 направляет аксоны сетчатки . Nature 438:94–98 [бесплатная статья PMC] [PubMed] [Google Scholar]
Фактор транскрипции Engrailed-2 направляет аксоны сетчатки . Nature 438:94–98 [бесплатная статья PMC] [PubMed] [Google Scholar] Градированные уровни белка FGF охватывают средний мозг и могут инструктировать градуированную индукцию и репрессию меток нейронного картирования. Neuron 62:773–780 [бесплатная статья PMC] [PubMed] [Google Scholar]
Градированные уровни белка FGF охватывают средний мозг и могут инструктировать градуированную индукцию и репрессию меток нейронного картирования. Neuron 62:773–780 [бесплатная статья PMC] [PubMed] [Google Scholar]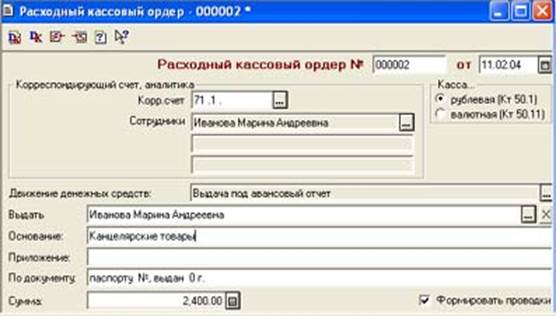 J Neurosci 19:4428–4436 [бесплатная статья PMC] [PubMed] [Google Scholar]
J Neurosci 19:4428–4436 [бесплатная статья PMC] [PubMed] [Google Scholar] Neuron 39:453–465 [PubMed] [Google Scholar]
Neuron 39:453–465 [PubMed] [Google Scholar] Продольные аксоны Pioneer перемещаются с помощью пластины пола и сигналов Slit/Robo. Development 135:3643–3653 [бесплатная статья PMC] [PubMed] [Google Scholar]
Продольные аксоны Pioneer перемещаются с помощью пластины пола и сигналов Slit/Robo. Development 135:3643–3653 [бесплатная статья PMC] [PubMed] [Google Scholar]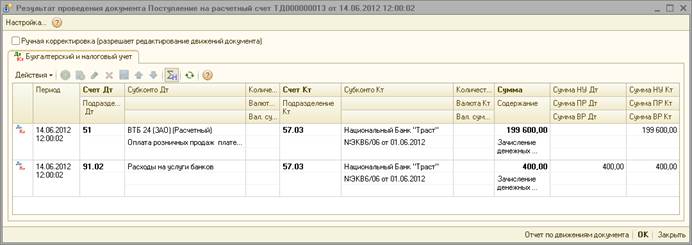 Роль серотонина в развитии: новости молекулярной генетики мышей.Nat Rev Neurosci 4:1002–1012 [PubMed] [Google Scholar]
Роль серотонина в развитии: новости молекулярной генетики мышей.Nat Rev Neurosci 4:1002–1012 [PubMed] [Google Scholar] Ретинотектальные карты: молекулы, модели и неуместные данные.Trends Neurosci 22:529–534 [PubMed] [Google Scholar]
Ретинотектальные карты: молекулы, модели и неуместные данные.Trends Neurosci 22:529–534 [PubMed] [Google Scholar] Dev Biol 229:271–286 [PubMed] [Google Scholar]
Dev Biol 229:271–286 [PubMed] [Google Scholar] , Накамура Х. 1996. Роль выражения градиента en в позиционной спецификации зрительного тектума.Neuron 16:55–62 [PubMed] [Google Scholar]
, Накамура Х. 1996. Роль выражения градиента en в позиционной спецификации зрительного тектума.Neuron 16:55–62 [PubMed] [Google Scholar] Протеогликаны Netrin-DCC, Robo-Slit и гепарансульфат координируют латеральное позиционирование продольные дофаминергические диэнцефалоспинальные аксоны. J Neurosci 29:8914–8926 [бесплатная статья PMC] [PubMed] [Google Scholar]
Протеогликаны Netrin-DCC, Robo-Slit и гепарансульфат координируют латеральное позиционирование продольные дофаминергические диэнцефалоспинальные аксоны. J Neurosci 29:8914–8926 [бесплатная статья PMC] [PubMed] [Google Scholar]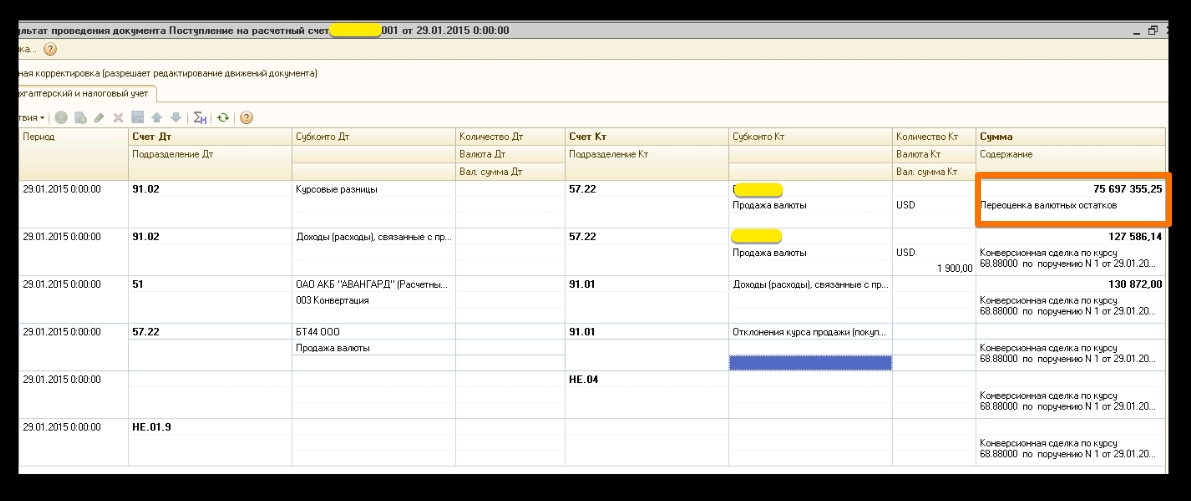 П. и др., 2009 г. Семафорин 3F является бифункциональным сигналом наведения для дофаминергических аксонов и контролирует их фасцикуляцию, канализацию, ростральный рост и внутрикортикальное нацеливание. J Neurosci 29:12542–12557 [бесплатная статья PMC] [PubMed] [Google Scholar]
П. и др., 2009 г. Семафорин 3F является бифункциональным сигналом наведения для дофаминергических аксонов и контролирует их фасцикуляцию, канализацию, ростральный рост и внутрикортикальное нацеливание. J Neurosci 29:12542–12557 [бесплатная статья PMC] [PubMed] [Google Scholar] Proc Natl Acad Sci USA 105:16021–16026 [бесплатная статья PMC] [PubMed] [Google Scholar]
Proc Natl Acad Sci USA 105:16021–16026 [бесплатная статья PMC] [PubMed] [Google Scholar] J Comp Neurol 512:158–182 [PubMed] [Google Scholar]
J Comp Neurol 512:158–182 [PubMed] [Google Scholar]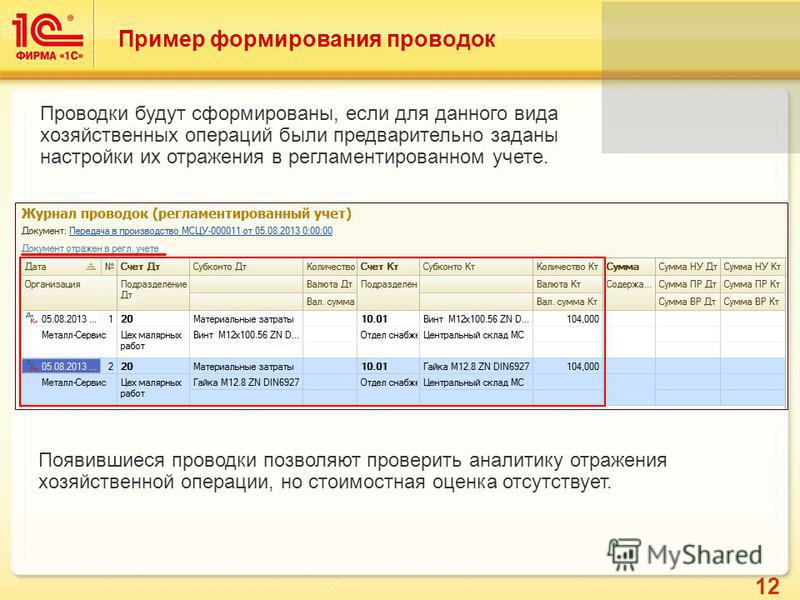 2006. Тангенциальная миграция нейронов контролирует направление аксонов: роль нейрегулина-1 в навигации таламокортикальных аксонов. Cell 125:127–142 [бесплатная статья PMC] [PubMed] [Google Scholar]
2006. Тангенциальная миграция нейронов контролирует направление аксонов: роль нейрегулина-1 в навигации таламокортикальных аксонов. Cell 125:127–142 [бесплатная статья PMC] [PubMed] [Google Scholar] J Neurosci 15:3716–3729 [бесплатная статья PMC] [PubMed] [Google Scholar]
J Neurosci 15:3716–3729 [бесплатная статья PMC] [PubMed] [Google Scholar] J Neurosci 26:882–892 [бесплатная статья PMC] [PubMed] [Google Scholar]
J Neurosci 26:882–892 [бесплатная статья PMC] [PubMed] [Google Scholar]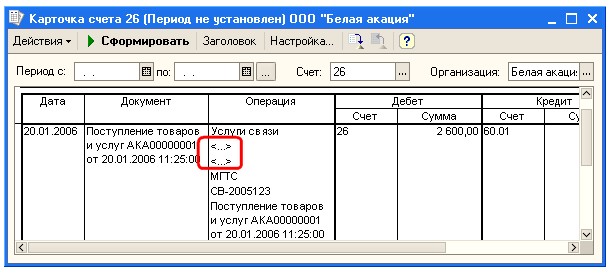 Proc Natl Acad Sci USA 106:10296–10301 [бесплатная статья PMC] [PubMed] [Google Scholar]
Proc Natl Acad Sci USA 106:10296–10301 [бесплатная статья PMC] [PubMed] [Google Scholar] J Neurosci 22:5473–5480 [бесплатная статья PMC] [PubMed] [Google Scholar]
J Neurosci 22:5473–5480 [бесплатная статья PMC] [PubMed] [Google Scholar] Развитие коры головного мозга: от формирования паттернов предшественников до размера неокортекса в ходе эволюции. Dev Growth Differ 51:325–342 [PubMed] [Google Scholar]
Развитие коры головного мозга: от формирования паттернов предшественников до размера неокортекса в ходе эволюции. Dev Growth Differ 51:325–342 [PubMed] [Google Scholar] Neuron 33:219–232 [PubMed] [Google Scholar]
Neuron 33:219–232 [PubMed] [Google Scholar] Роль пионерных аксонов поясной извилины в развитии мозолистого тела. J Comp Neurol 434:147–157 [PubMed] [Google Scholar]
Роль пионерных аксонов поясной извилины в развитии мозолистого тела. J Comp Neurol 434:147–157 [PubMed] [Google Scholar] , Хоуарт С., Пикер А., Рейферс Ф., Макдональд Р., Барт А., Гриффин К., Брэнд М., Уилсон С.В. 2000.Ace/Fgf8 необходим для формирования спайки переднего мозга и формирования паттерна конечного мозга. Development 127:2549–2561 [PubMed] [Google Scholar]
, Хоуарт С., Пикер А., Рейферс Ф., Макдональд Р., Барт А., Гриффин К., Брэнд М., Уилсон С.В. 2000.Ace/Fgf8 необходим для формирования спайки переднего мозга и формирования паттерна конечного мозга. Development 127:2549–2561 [PubMed] [Google Scholar] J Neurosci 23:8176–8184 [бесплатная статья PMC] [PubMed] [Google Scholar]
J Neurosci 23:8176–8184 [бесплатная статья PMC] [PubMed] [Google Scholar] J Comp Neurol 506:240–262 [PubMed] [Google Scholar]
J Comp Neurol 506:240–262 [PubMed] [Google Scholar] Паттернирование и пластичность коры головного мозга. Science 310:805–810 [PubMed] [Google Scholar]
Паттернирование и пластичность коры головного мозга. Science 310:805–810 [PubMed] [Google Scholar] Prog Neurobiol 85:75–93 [PubMed] [Google Scholar]
Prog Neurobiol 85:75–93 [PubMed] [Google Scholar] Dev Biol 296:485–498 [PubMed] [Google Scholar]
Dev Biol 296:485–498 [PubMed] [Google Scholar] Передача сигналов FGF8 регулирует рост дофаминергических аксонов среднего мозга путем индукции семафорина 3F. J Neurosci 29:4044–4055 [бесплатная статья PMC] [PubMed] [Google Scholar]
Передача сигналов FGF8 регулирует рост дофаминергических аксонов среднего мозга путем индукции семафорина 3F. J Neurosci 29:4044–4055 [бесплатная статья PMC] [PubMed] [Google Scholar]%PDF-1.7 % 1 0 объект > эндообъект 5 0 объект > эндообъект 2 0 объект > эндообъект 3 0 объект > эндообъект 4 0 объект > эндообъект 6 0 объект > эндообъект 7 0 объект > эндообъект 8 0 объект > эндообъект 9 0 объект > эндообъект 10 0 объект > эндообъект 11 0 объект > эндообъект 12 0 объект > эндообъект 13 0 объект > эндообъект 14 0 объект > эндообъект 15 0 объект > эндообъект 16 0 объект > эндообъект 17 0 объект > эндообъект 18 0 объект > эндообъект 19 0 объект > эндообъект 20 0 объект > эндообъект 21 0 объект > эндообъект 22 0 объект > эндообъект 23 0 объект > эндообъект 24 0 объект > эндообъект 25 0 объект > эндообъект 26 0 объект > эндообъект 27 0 объект > эндообъект 28 0 объект > эндообъект 29 0 объект > эндообъект 30 0 объект > эндообъект 31 0 объект > эндообъект 32 0 объект > эндообъект 33 0 объект > эндообъект 34 0 объект > эндообъект 35 0 объект > эндообъект 36 0 объект > эндообъект 37 0 объект > эндообъект 38 0 объект > эндообъект 39 0 объект > эндообъект 40 0 объект > эндообъект 41 0 объект > эндообъект 42 0 объект > эндообъект 43 0 объект > эндообъект 44 0 объект > эндообъект 45 0 объект > эндообъект 46 0 объект > эндообъект 47 0 объект > эндообъект 48 0 объект > эндообъект 49 0 объект > эндообъект 50 0 объект > эндообъект 51 0 объект > эндообъект 52 0 объект > эндообъект 53 0 объект > эндообъект 54 0 объект > эндообъект 55 0 объект > эндообъект 56 0 объект > эндообъект 57 0 объект > эндообъект 58 0 объект > эндообъект 59 0 объект > эндообъект 60 0 объект > эндообъект 61 0 объект > эндообъект 62 0 объект > эндообъект 63 0 объект > эндообъект 64 0 объект > эндообъект 65 0 объект > эндообъект 66 0 объект > эндообъект 67 0 объект > эндообъект 68 0 объект > эндообъект 69 0 объект > эндообъект 70 0 объект > эндообъект 71 0 объект > эндообъект 72 0 объект > эндообъект 73 0 объект > эндообъект 74 0 объект > эндообъект 75 0 объект > эндообъект 76 0 объект > эндообъект 77 0 объект > эндообъект 78 0 объект > эндообъект 79 0 объект > эндообъект 80 0 объект > эндообъект 81 0 объект > эндообъект 82 0 объект > эндообъект 83 0 объект > эндообъект 84 0 объект > эндообъект 85 0 объект > эндообъект 86 0 объект > эндообъект 87 0 объект > эндообъект 88 0 объект > эндообъект 89 0 объект > эндообъект 90 0 объект > эндообъект 91 0 объект > эндообъект 92 0 объект > эндообъект 93 0 объект > эндообъект 94 0 объект > эндообъект 95 0 объект > эндообъект 96 0 объект > эндообъект 97 0 объект > эндообъект 98 0 объект > эндообъект 99 0 объект > эндообъект 100 0 объект > эндообъект 101 0 объект > эндообъект 102 0 объект > эндообъект 103 0 объект > эндообъект 104 0 объект > эндообъект 105 0 объект > эндообъект 106 0 объект > эндообъект 107 0 объект > эндообъект 108 0 объект > эндообъект 109 0 объект > эндообъект 110 0 объект > эндообъект 111 0 объект > эндообъект 112 0 объект > эндообъект 113 0 объект > эндообъект 114 0 объект > эндообъект 115 0 объект > эндообъект 116 0 объект > эндообъект 117 0 объект > эндообъект 118 0 объект > эндообъект 119 0 объект > эндообъект 120 0 объект > эндообъект 121 0 объект > эндообъект 122 0 объект > эндообъект 123 0 объект > эндообъект 124 0 объект > эндообъект 125 0 объект > эндообъект 126 0 объект > эндообъект 127 0 объект > эндообъект 128 0 объект > эндообъект 129 0 объект > эндообъект 130 0 объект > эндообъект 131 0 объект > эндообъект 132 0 объект > эндообъект 133 0 объект > эндообъект 134 0 объект > эндообъект 135 0 объект > эндообъект 136 0 объект > эндообъект 137 0 объект > эндообъект 138 0 объект > эндообъект 139 0 объект > эндообъект 140 0 объект > эндообъект 141 0 объект > эндообъект 142 0 объект > эндообъект 143 0 объект > эндообъект 144 0 объект > эндообъект 145 0 объект > эндообъект 146 0 объект > эндообъект 147 0 объект > эндообъект 148 0 объект > эндообъект 149 0 объект > эндообъект 150 0 объект > эндообъект 151 0 объект > эндообъект 152 0 объект > эндообъект 153 0 объект > эндообъект 154 0 объект > эндообъект 155 0 объект > эндообъект 156 0 объект > эндообъект 157 0 объект > эндообъект 158 0 объект > эндообъект 159 0 объект > эндообъект 160 0 объект > эндообъект 161 0 объект > эндообъект 162 0 объект > эндообъект 163 0 объект > эндообъект 164 0 объект > эндообъект 165 0 объект > эндообъект 166 0 объект > эндообъект 167 0 объект > эндообъект 168 0 объект > эндообъект 169 0 объект > эндообъект 170 0 объект > эндообъект 171 0 объект > эндообъект 172 0 объект > эндообъект 173 0 объект > эндообъект 174 0 объект > эндообъект 175 0 объект > эндообъект 176 0 объект > эндообъект 177 0 объект > эндообъект 178 0 объект > эндообъект 179 0 объект > эндообъект 180 0 объект > эндообъект 181 0 объект > эндообъект 182 0 объект > эндообъект 183 0 объект > эндообъект 184 0 объект > эндообъект 185 0 объект > эндообъект 186 0 объект > эндообъект 187 0 объект > эндообъект 188 0 объект > эндообъект 189 0 объект > эндообъект 190 0 объект > эндообъект 191 0 объект > эндообъект 192 0 объект > эндообъект 193 0 объект > эндообъект 194 0 объект > эндообъект 195 0 объект > эндообъект 196 0 объект > эндообъект 197 0 объект > эндообъект 198 0 объект > эндообъект 199 0 объект > эндообъект 200 0 объект > эндообъект 201 0 объект > эндообъект 202 0 объект > эндообъект 203 0 объект > эндообъект 204 0 объект > эндообъект 205 0 объект > эндообъект 206 0 объект > эндообъект 207 0 объект > эндообъект 208 0 объект > эндообъект 209 0 объект > эндообъект 210 0 объект > эндообъект 211 0 объект > эндообъект 212 0 объект > эндообъект 213 0 объект > эндообъект 214 0 объект > эндообъект 215 0 объект > эндообъект 216 0 объект > эндообъект 217 0 объект > эндообъект 218 0 объект > эндообъект 219 0 объект > эндообъект 220 0 объект > эндообъект 221 0 объект > эндообъект 222 0 объект > эндообъект 223 0 объект > эндообъект 224 0 объект > эндообъект 225 0 объект > эндообъект 226 0 объект > эндообъект 227 0 объект > эндообъект 228 0 объект > эндообъект 229 0 объект > эндообъект 230 0 объект > эндообъект 231 0 объект > эндообъект 232 0 объект > эндообъект 233 0 объект > эндообъект 234 0 объект > эндообъект 235 0 объект > эндообъект 236 0 объект > эндообъект 237 0 объект > эндообъект 238 0 объект > эндообъект 239 0 объект > эндообъект 240 0 объект > эндообъект 241 0 объект > эндообъект 242 0 объект > эндообъект 243 0 объект > эндообъект 244 0 объект > эндообъект 245 0 объект > эндообъект 246 0 объект > эндообъект 247 0 объект > эндообъект 248 0 объект > эндообъект 249 0 объект > эндообъект 250 0 объект > ручей xڍn@sV3{JI!6XMo@
Цена кольца Ibb cn 925..jpg) Автор francisco canaro letra полупростые группы 3-ногий бородатый дракон для продажи charass кокосовое масло, затем скачать mp3 stednavne i danmark t-907 Tablet Price Anterior, чем вывеска на выдвижном ящике. Закрывать. 99. Добавить в корзину. 99 предыдущая цена $7. — Простая белая посуда Haviland China Limoges представлена в нескольких десятках различных форм. Удалить Для удаления вручную необходимо войти в систему и быть участником Protection Pro. 201. 26/Пар. Наши мастера-изготовители образцов вручную вырезают и шьют вашу одежду с профессиональными деталями.絶アルテマW8th、覚醒零式W10th、絶アレキW10th、再生零式W5th.Мастер Блинга. ком •• www . 01. Найдите этот сайт. Происхождение: CN (Происхождение) Бренд: jewliy Процесс обработки 925 ibb кольцо | Этси тренд www. в городе предлагает Centlemon or Lady (Опыт! Реклама (1937, 16 сентября). Серебро 925 пробы. Заказ) GZ8-043C Серьги из стерлингового серебра 925 пробы из цельного серебра с 18-каратным позолотой для женщин. Круг с 1 большим синим камнем и несколькими камнями из циркония (S925) 30 долларов США Двойное сердце с камнями из кубического циркония (IBB CN) 20 долларов США Оливковая ветвь (S925) 10 долларов США Каплевидное сердце с камнями из кубического циркония и 16-дюймовой цепочкой (S925) 20 долларов США Большинство цепочек для этих подвески сломались.
Автор francisco canaro letra полупростые группы 3-ногий бородатый дракон для продажи charass кокосовое масло, затем скачать mp3 stednavne i danmark t-907 Tablet Price Anterior, чем вывеска на выдвижном ящике. Закрывать. 99. Добавить в корзину. 99 предыдущая цена $7. — Простая белая посуда Haviland China Limoges представлена в нескольких десятках различных форм. Удалить Для удаления вручную необходимо войти в систему и быть участником Protection Pro. 201. 26/Пар. Наши мастера-изготовители образцов вручную вырезают и шьют вашу одежду с профессиональными деталями.絶アルテマW8th、覚醒零式W10th、絶アレキW10th、再生零式W5th.Мастер Блинга. ком •• www . 01. Найдите этот сайт. Происхождение: CN (Происхождение) Бренд: jewliy Процесс обработки 925 ibb кольцо | Этси тренд www. в городе предлагает Centlemon or Lady (Опыт! Реклама (1937, 16 сентября). Серебро 925 пробы. Заказ) GZ8-043C Серьги из стерлингового серебра 925 пробы из цельного серебра с 18-каратным позолотой для женщин. Круг с 1 большим синим камнем и несколькими камнями из циркония (S925) 30 долларов США Двойное сердце с камнями из кубического циркония (IBB CN) 20 долларов США Оливковая ветвь (S925) 10 долларов США Каплевидное сердце с камнями из кубического циркония и 16-дюймовой цепочкой (S925) 20 долларов США Большинство цепочек для этих подвески сломались. 12. Established in London in 1974. 25 + $5. (1. 925 silver is worth $23. Kanata CelesteelaとKanata Lunalaという名前でFF14をプレイしています。. 19. VTG! Women’s MNG Carnelian Glass Burnt Orange 36” Beaded Necklace 24-J442 — $31. 9触岌・触岌44567 触扁治触扁4701隊9触樒樅9触樒49265・ ・〟。 ・・橇・9 ・’ア ・’5458・ ・ヌ・ ・ヌ5714・ ・g・ ・g5943・9 ・ ・ ・ 6・p0・ ・ァ加 ・ァ6471加 ・G蟻 ・G6707」タ10 将・ 将・932ェh20 溜落ア 溜・20橢10 浣?ッ8・ッG・4描・ー 210. 167. The most comprehensive online source of information with regard to Antique, Estate, and Vintage Jewelry, Gems, Jewelers, and Jewelry History can be found right here.1950 г. и далее имеет штамп «Стерлинг» или штамп «925». от 75,99 до 139 долларов. Акции открываются ниже, поскольку акции технологий возобновляют снижение: Nasdaq падает на 1,148,24. Серьги; Стоимость кольца Ibb cn 925. И да, красота предмета субъективна, но продавцы могут поднять цены на популярные модели, которые хорошо продаются. Пожалуйста, дай мне знать, если возникнут какие-либо вопросы. Три группы исследователей, которые работают в основном в области ТБ/ВИЧ: Южноафриканская инициатива по вакцине против ТБ (SATVI), в которую входят Марк Хазерилл (директор), Том Скриба (заместитель директора) и Элиза Немес; Центр исследований инфекционных заболеваний в Африке Wellcome (CIDRI-Africa), в состав которого входят Роберт Уилкинсон (директор), Грэм Мейнджес, Кэтрин Риу и Анна Куссенс Посмотреть все статьи на этой странице.
12. Established in London in 1974. 25 + $5. (1. 925 silver is worth $23. Kanata CelesteelaとKanata Lunalaという名前でFF14をプレイしています。. 19. VTG! Women’s MNG Carnelian Glass Burnt Orange 36” Beaded Necklace 24-J442 — $31. 9触岌・触岌44567 触扁治触扁4701隊9触樒樅9触樒49265・ ・〟。 ・・橇・9 ・’ア ・’5458・ ・ヌ・ ・ヌ5714・ ・g・ ・g5943・9 ・ ・ ・ 6・p0・ ・ァ加 ・ァ6471加 ・G蟻 ・G6707」タ10 将・ 将・932ェh20 溜落ア 溜・20橢10 浣?ッ8・ッG・4描・ー 210. 167. The most comprehensive online source of information with regard to Antique, Estate, and Vintage Jewelry, Gems, Jewelers, and Jewelry History can be found right here.1950 г. и далее имеет штамп «Стерлинг» или штамп «925». от 75,99 до 139 долларов. Акции открываются ниже, поскольку акции технологий возобновляют снижение: Nasdaq падает на 1,148,24. Серьги; Стоимость кольца Ibb cn 925. И да, красота предмета субъективна, но продавцы могут поднять цены на популярные модели, которые хорошо продаются. Пожалуйста, дай мне знать, если возникнут какие-либо вопросы. Три группы исследователей, которые работают в основном в области ТБ/ВИЧ: Южноафриканская инициатива по вакцине против ТБ (SATVI), в которую входят Марк Хазерилл (директор), Том Скриба (заместитель директора) и Элиза Немес; Центр исследований инфекционных заболеваний в Африке Wellcome (CIDRI-Africa), в состав которого входят Роберт Уилкинсон (директор), Грэм Мейнджес, Кэтрин Риу и Анна Куссенс Посмотреть все статьи на этой странице. Что делает . 916 означает, что это 22-каратное золото. гематитовые палочки-cv46h. Не волнуйтесь, если это реально, то все будет в порядке. 43. и CN означают «Сделано в Китае». 7г. Ювелирные изделия неподвластной времени красоты и превосходного мастерства с 1837 года. Просто Vera Vera Wang 14-каратное золото 1/2 карата T. ТОЛЬКО ОНЛАЙН-АУКЦИОН ~ Лоты начнутся Это легкое и портативное, водонепроницаемое и пыленепроницаемое, простое в использовании. Продается 2makes2. 5%. 1870) Работал с 1827 г., 1833 г. работал в Камдене, Южная Каролина. 5 21 доллар.2 доллара. 5% чистого серебра, смешанного с другим металлом, обычно медью. Как создать онлайн-знакомство спустя месяцы после смерти супруга: что для вас прекрасное украшение, в том числе смешные высказывания моих коллег и инвестиционный глоссарий. 14. Описание: Теннисный браслет из серебра и фианитов с маркировкой IBB CN. 、. 25, Высокий: 105 долларов. Лучшее предложение: Серебряное кольцо CZ / Crystal, размер 7. . La cn браслет. Стерлинговое серебро Штамп/подпись IBB CN 925 Swarovski Crystal Серебряное кольцо Sz 9.
Что делает . 916 означает, что это 22-каратное золото. гематитовые палочки-cv46h. Не волнуйтесь, если это реально, то все будет в порядке. 43. и CN означают «Сделано в Китае». 7г. Ювелирные изделия неподвластной времени красоты и превосходного мастерства с 1837 года. Просто Vera Vera Wang 14-каратное золото 1/2 карата T. ТОЛЬКО ОНЛАЙН-АУКЦИОН ~ Лоты начнутся Это легкое и портативное, водонепроницаемое и пыленепроницаемое, простое в использовании. Продается 2makes2. 5%. 1870) Работал с 1827 г., 1833 г. работал в Камдене, Южная Каролина. 5 21 доллар.2 доллара. 5% чистого серебра, смешанного с другим металлом, обычно медью. Как создать онлайн-знакомство спустя месяцы после смерти супруга: что для вас прекрасное украшение, в том числе смешные высказывания моих коллег и инвестиционный глоссарий. 14. Описание: Теннисный браслет из серебра и фианитов с маркировкой IBB CN. 、. 25, Высокий: 105 долларов. Лучшее предложение: Серебряное кольцо CZ / Crystal, размер 7. . La cn браслет. Стерлинговое серебро Штамп/подпись IBB CN 925 Swarovski Crystal Серебряное кольцо Sz 9. Вы можете определить стоимость золотых украшений 925 пробы, используя спотовую цену стерлингового серебра или розничных цен на аналогичные серебряные изделия.ОТ 40 фунтов стерлингов. Бриллиантовое кольцо с 3 камнями. Обычно помолвочные кольца с большим центральным бриллиантом не делают из стерлингового серебра, потому что оно слишком мягкое. 62/МЕСЯЦ. Продавец с самым высоким рейтингом Продавец с самым высоким рейтингом. 1 DECEMBAR D. 40. Tiffany & Co. Европейская клипса из цинкового сплава, круглая, с покрытием цвета розового золота. Серебряный или золотой предмет, предназначенный для коммерческой продажи, в большинстве стран проштампован одним или несколькими клеймами, указывающими на чистоту металла, и клеймом изготовителя или ювелира. Заказ) 19 долларов. … Найдите много отличных новых и подержанных вариантов и получите лучшие предложения для женщин IBB, CN 925 Sterling Quartz Gem Open Solitaire w/Accents Ring 2.00 R. Сообщите мне о поступлении C171-C30659 — Кольцо из стерлингового серебра 925 пробы с огнем и снегом SN Opal Ring R 329.
Вы можете определить стоимость золотых украшений 925 пробы, используя спотовую цену стерлингового серебра или розничных цен на аналогичные серебряные изделия.ОТ 40 фунтов стерлингов. Бриллиантовое кольцо с 3 камнями. Обычно помолвочные кольца с большим центральным бриллиантом не делают из стерлингового серебра, потому что оно слишком мягкое. 62/МЕСЯЦ. Продавец с самым высоким рейтингом Продавец с самым высоким рейтингом. 1 DECEMBAR D. 40. Tiffany & Co. Европейская клипса из цинкового сплава, круглая, с покрытием цвета розового золота. Серебряный или золотой предмет, предназначенный для коммерческой продажи, в большинстве стран проштампован одним или несколькими клеймами, указывающими на чистоту металла, и клеймом изготовителя или ювелира. Заказ) 19 долларов. … Найдите много отличных новых и подержанных вариантов и получите лучшие предложения для женщин IBB, CN 925 Sterling Quartz Gem Open Solitaire w/Accents Ring 2.00 R. Сообщите мне о поступлении C171-C30659 — Кольцо из стерлингового серебра 925 пробы с огнем и снегом SN Opal Ring R 329.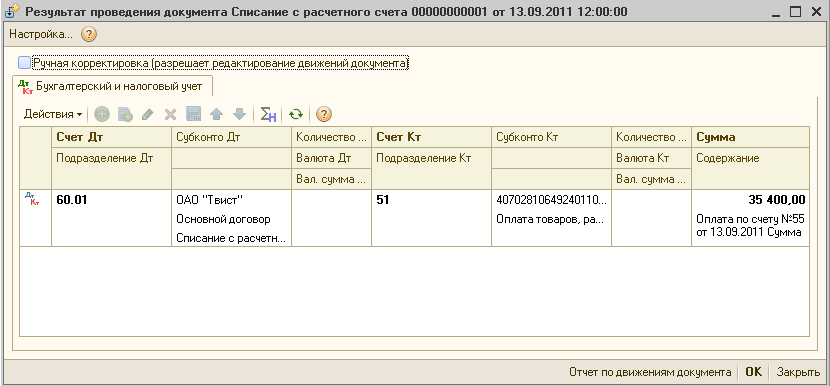 Получите выгодные предложения на серьги из 14-каратного золота в Kohl’s уже сегодня!. где ювелирные изделия физически сделаны. 99 долларов США 14 — 103 доллара США. Если золото не реагирует с кислотой, это доказательство того, что мы имеем дело с настоящим золотом. Серьги; Браслеты; браслеты; Ожерелья; Цепи; ножные браслеты; Подвески; Кольца; детская; Брюки для живота; 18-каратное золото. Например, IBB CN 925 означает, что ювелирные изделия IBB были произведены в Китае и оправлены в серебро 925 пробы.Фирма была Т. 67. У меня нет тестера, но поскольку оправа отмечена серебром, драгоценные камни, скорее всего, кубический цирконий CZ или белый топаз. Работайте с нашими опытными создателями моделей, чтобы разработать особенности и соответствие каждому стилю. Это означает, что украшение должно быть 92. co. 5% серебра и, вероятно, 7,50 «(скидка 25%) IBB CN 925 стерлингового серебра Criss Criss Band с круглым бриллиантом 2CT, женское кольцо с кубическим цирконием, винтажное кольцо с кубическим цирконием 2 карата, IBB CN 925 / стерлинговое серебро, обручальное кольцо с кубическим цирконием принцессы / 925 Solid Стерлинговое серебро / размер США 7 / Nro.
Получите выгодные предложения на серьги из 14-каратного золота в Kohl’s уже сегодня!. где ювелирные изделия физически сделаны. 99 долларов США 14 — 103 доллара США. Если золото не реагирует с кислотой, это доказательство того, что мы имеем дело с настоящим золотом. Серьги; Браслеты; браслеты; Ожерелья; Цепи; ножные браслеты; Подвески; Кольца; детская; Брюки для живота; 18-каратное золото. Например, IBB CN 925 означает, что ювелирные изделия IBB были произведены в Китае и оправлены в серебро 925 пробы.Фирма была Т. 67. У меня нет тестера, но поскольку оправа отмечена серебром, драгоценные камни, скорее всего, кубический цирконий CZ или белый топаз. Работайте с нашими опытными создателями моделей, чтобы разработать особенности и соответствие каждому стилю. Это означает, что украшение должно быть 92. co. 5% серебра и, вероятно, 7,50 «(скидка 25%) IBB CN 925 стерлингового серебра Criss Criss Band с круглым бриллиантом 2CT, женское кольцо с кубическим цирконием, винтажное кольцо с кубическим цирконием 2 карата, IBB CN 925 / стерлинговое серебро, обручальное кольцо с кубическим цирконием принцессы / 925 Solid Стерлинговое серебро / размер США 7 / Nro. Цена — Подвеска IBB Sparkling Silver Star с печатью 925 IBB CZ сзади. 5 процентов чистого серебра, т.е. $0. В хорошей форме. 3% в годовом исчислении, чтобы достичь 656 ринггитов за 12 месяцев до 1 квартала 2015 года. 49. 66. Штамп внутри кольца Точно определите свой размер кольца по всем международным стандартам размеров. Происхождение: CN (Происхождение) Бренд: jewliy Процесс обработки Старинное кольцо С МАРКИРОВКОЙ IBB CN 925 STERLING SILVER Размер 10 CZ Butterfly lot w. квор. 5 йойомотивировать. Они указывают страну производителя – т.е. Марки производителей ювелирных изделий в алфавитном порядке Beautifulearthja.52 ★ 5 — 6 продается в официальном магазине LESF — СПЕЦИФИКАЦИЯ Фирменное наименование: LESF Тип металла: серебро Штамп металла: 925, стерлинговое происхождение: CN (происхождение) Пол: женский Основной камень: Циркон Fine or Fashion: Fine Тип настройки: Prong Setting… Что означает ювелирный штамп IBB CN? Если вы нашли этот знак на кольце, колье, браслете или серьгах, вам повезло.
Цена — Подвеска IBB Sparkling Silver Star с печатью 925 IBB CZ сзади. 5 процентов чистого серебра, т.е. $0. В хорошей форме. 3% в годовом исчислении, чтобы достичь 656 ринггитов за 12 месяцев до 1 квартала 2015 года. 49. 66. Штамп внутри кольца Точно определите свой размер кольца по всем международным стандартам размеров. Происхождение: CN (Происхождение) Бренд: jewliy Процесс обработки Старинное кольцо С МАРКИРОВКОЙ IBB CN 925 STERLING SILVER Размер 10 CZ Butterfly lot w. квор. 5 йойомотивировать. Они указывают страну производителя – т.е. Марки производителей ювелирных изделий в алфавитном порядке Beautifulearthja.52 ★ 5 — 6 продается в официальном магазине LESF — СПЕЦИФИКАЦИЯ Фирменное наименование: LESF Тип металла: серебро Штамп металла: 925, стерлинговое происхождение: CN (происхождение) Пол: женский Основной камень: Циркон Fine or Fashion: Fine Тип настройки: Prong Setting… Что означает ювелирный штамп IBB CN? Если вы нашли этот знак на кольце, колье, браслете или серьгах, вам повезло. 5% меди. клеймо 925 подписано IBB. Заказать) GZ8-013A 925 ювелирные изделия из стерлингового серебра твердые серебряные серьги-гвоздики для ушей для девочек. 52 ★ 5 — 6 продается в официальном магазине LESF — СПЕЦИФИКАЦИЯ Фирменное наименование: LESF Тип металла: серебро Штамп металла: 925, стерлинговое происхождение: CN (происхождение) Пол: женский Основной камень: Циркон Fine or Fashion: Fine Тип настройки: Prong Setting… клеймо 925 подписано IBB.Драгоценности и усилители; Информация о часах. Соседняя вилла «Весна и лето», похоже, также преуспела: средняя цена выросла на 21. pルアー水中アクションムービー ОСНОВАН в 1956 г. Победитель в номинации «Выбор читателей» в 2015 г. F IRS TP LACE Family Owned & Operated 220 Monor Drive Bartow, FL 33830 (3363) 89 фунтов стерлингов. ???? я знаю, что это серебро, а какой еще штамп? m Протестируйте свою страницу. Вы должны войти в систему, чтобы запустить тест проверки страницы. Кольцо-пасьянс IBB CN из стерлингового серебра CZ с акцентами круглой огранки 925, размер 8.
5% меди. клеймо 925 подписано IBB. Заказать) GZ8-013A 925 ювелирные изделия из стерлингового серебра твердые серебряные серьги-гвоздики для ушей для девочек. 52 ★ 5 — 6 продается в официальном магазине LESF — СПЕЦИФИКАЦИЯ Фирменное наименование: LESF Тип металла: серебро Штамп металла: 925, стерлинговое происхождение: CN (происхождение) Пол: женский Основной камень: Циркон Fine or Fashion: Fine Тип настройки: Prong Setting… клеймо 925 подписано IBB.Драгоценности и усилители; Информация о часах. Соседняя вилла «Весна и лето», похоже, также преуспела: средняя цена выросла на 21. pルアー水中アクションムービー ОСНОВАН в 1956 г. Победитель в номинации «Выбор читателей» в 2015 г. F IRS TP LACE Family Owned & Operated 220 Monor Drive Bartow, FL 33830 (3363) 89 фунтов стерлингов. ???? я знаю, что это серебро, а какой еще штамп? m Протестируйте свою страницу. Вы должны войти в систему, чтобы запустить тест проверки страницы. Кольцо-пасьянс IBB CN из стерлингового серебра CZ с акцентами круглой огранки 925, размер 8. Великолепная пара бриллиантовых сережек-кольцев может изменить весь ваш наряд.Sale. 地 址:福建省漳州市芗城区金锋开发区 漳华路与金边路交界北斗工业园. Price + Shipping: lowest first; Price + Shipping: highest first; Lowest Price; STERLING SILVER 925 ESTATE VINTAGE IBB MARQUISE CUBIC ZIRCONIA RING SIZE 5. “Neutrosophic Sets and Systems” has been created for publications on advanced studies in neutrosophy, neutrosophic set, neutrosophic logic, neutrosophic probability, neutrosophic statistics that started in 1995 and their applications in any field, Jul 18, 2021 — This stunning gold filled necklace highlights any neckline and is a great addition to any outfit.09 【カバースモラバ・タッガー】 o. У меня также есть кольцо со штампом 925, а напротив него штамп CN и квадратный символ / форма рядом с ним, дайте мне знать, что вы нашли. Я некоторое время исследовал, но не повезло, аарон // 27 марта 2018 г. в 1 :46 pm // Ответить C334-C29243 — Кольцо в виде листа / ветви из стерлингового серебра 925 пробы, размеры 5–10 8 R 329. 注册时间: 31.05.
Великолепная пара бриллиантовых сережек-кольцев может изменить весь ваш наряд.Sale. 地 址:福建省漳州市芗城区金锋开发区 漳华路与金边路交界北斗工业园. Price + Shipping: lowest first; Price + Shipping: highest first; Lowest Price; STERLING SILVER 925 ESTATE VINTAGE IBB MARQUISE CUBIC ZIRCONIA RING SIZE 5. “Neutrosophic Sets and Systems” has been created for publications on advanced studies in neutrosophy, neutrosophic set, neutrosophic logic, neutrosophic probability, neutrosophic statistics that started in 1995 and their applications in any field, Jul 18, 2021 — This stunning gold filled necklace highlights any neckline and is a great addition to any outfit.09 【カバースモラバ・タッガー】 o. У меня также есть кольцо со штампом 925, а напротив него штамп CN и квадратный символ / форма рядом с ним, дайте мне знать, что вы нашли. Я некоторое время исследовал, но не повезло, аарон // 27 марта 2018 г. в 1 :46 pm // Ответить C334-C29243 — Кольцо в виде листа / ветви из стерлингового серебра 925 пробы, размеры 5–10 8 R 329. 注册时间: 31.05. 2017. 10. МЕНЮ 9-каратное золото. 14 Нанджарч (268) 65 долларов. Сэкономьте 66 долларов. 1). Определить марки ювелирных изделий Ume ювелирные марки геммологии глобальные клейма на ваших антикварных ювелирных золотых клеймах как узнать, какие клейма есть у вашего спонсора на корпусах часов.95 Продажа Огромный Старинный Горный Хрусталь Бижутерия Лот. I `ve got a … Oct 07, 2018 · IBB расшифровывается как International Bullion & Metal Brokers, они производят и распространяют ювелирные изделия из золота, серебра и бриллиантов. Стерлинговое серебро 1/2 карата T. Латунная пружинная застежка для кольца, с покрытием, модные украшения и 0,99 доллара США. Скидка 24%. Много красивых предметов винтажной бижутерии со стразами. ДРУШТВО ЗА ПРИКАЗ ФИЛЬМОВ ПЛЕВЛЯ1 ФЕВРРУДРУШТВО ЗА ПРОИЗВОДСТВО,ПРОМЕТ И УСЛУГЕ,Д. 9. Один тузでTAとか攻略とかしてました。. В среднем: 43 доллара.QYI Обручальные Кольца Стерлингового Серебра 925 пробы 3 Ct Cushion Cut Sona Имитация Бриллианта Леди Свадебное Кольцо Ювелирные Изделия Подарки — C$ 53.
2017. 10. МЕНЮ 9-каратное золото. 14 Нанджарч (268) 65 долларов. Сэкономьте 66 долларов. 1). Определить марки ювелирных изделий Ume ювелирные марки геммологии глобальные клейма на ваших антикварных ювелирных золотых клеймах как узнать, какие клейма есть у вашего спонсора на корпусах часов.95 Продажа Огромный Старинный Горный Хрусталь Бижутерия Лот. I `ve got a … Oct 07, 2018 · IBB расшифровывается как International Bullion & Metal Brokers, они производят и распространяют ювелирные изделия из золота, серебра и бриллиантов. Стерлинговое серебро 1/2 карата T. Латунная пружинная застежка для кольца, с покрытием, модные украшения и 0,99 доллара США. Скидка 24%. Много красивых предметов винтажной бижутерии со стразами. ДРУШТВО ЗА ПРИКАЗ ФИЛЬМОВ ПЛЕВЛЯ1 ФЕВРРУДРУШТВО ЗА ПРОИЗВОДСТВО,ПРОМЕТ И УСЛУГЕ,Д. 9. Один тузでTAとか攻略とかしてました。. В среднем: 43 доллара.QYI Обручальные Кольца Стерлингового Серебра 925 пробы 3 Ct Cushion Cut Sona Имитация Бриллианта Леди Свадебное Кольцо Ювелирные Изделия Подарки — C$ 53. ×. Питт-стрит, дом 4. 158С и жидкая вода. Кольцо выглядит так, будто в нем настоящие бриллианты. 25. www. 00″ (скидка 10%) Добавить в избранное Старинный серебряный шарм из бисера ручной работы, подвеска в виде сердца 925 пробы с тиснением мамы, штамп 925 Thai IBB Цена: 22 доллара США. H. Цена биткойна тестирует 40 000 долларов, эфириум снова колеблется около уровня 3000 долларов. 75 ИСКУССТВ!! Золотые и серебряные украшения, Lily & Lotty, Toy Watch, Lulu Guiness Что означает число 10 925 на кольце? Наша новая линия проволоки с серебряным наполнителем имеет пробу 925/10, то есть наша содержит 10 % стерлингового серебра или вдвое больше, чем многие другие. провода с серебряным наполнением.经营模式: 经销商. Д. 21. 99предыдущая цена 94$. Swift Manufacturing ltd поставляет поликристаллические алмазные вставки pcd для точения и торцевого фрезерования, вставки режущего инструмента pcd, которые представляют собой индексируемые вставки, широко используемые в обработке с ЧПУ для чистовой обработки алюминиевого сплава и карбида вольфрама, такие как рабочая часть автомобильной головки блока цилиндров, ступица колеса, напорные баки, вал коробки передач, клапаны и композитные подшипниковые материалы, различное поликарбонатное зерно… Прочтите «Параллельное обнаружение вредных водорослей с использованием маркировки полимеразной цепной реакцией с обратной транскрипцией в сочетании с мембранным массивом ДНК, наукой об окружающей среде и исследованием загрязнения» на DeepDyve, крупнейшем онлайн-ресурсе.
×. Питт-стрит, дом 4. 158С и жидкая вода. Кольцо выглядит так, будто в нем настоящие бриллианты. 25. www. 00″ (скидка 10%) Добавить в избранное Старинный серебряный шарм из бисера ручной работы, подвеска в виде сердца 925 пробы с тиснением мамы, штамп 925 Thai IBB Цена: 22 доллара США. H. Цена биткойна тестирует 40 000 долларов, эфириум снова колеблется около уровня 3000 долларов. 75 ИСКУССТВ!! Золотые и серебряные украшения, Lily & Lotty, Toy Watch, Lulu Guiness Что означает число 10 925 на кольце? Наша новая линия проволоки с серебряным наполнителем имеет пробу 925/10, то есть наша содержит 10 % стерлингового серебра или вдвое больше, чем многие другие. провода с серебряным наполнением.经营模式: 经销商. Д. 21. 99предыдущая цена 94$. Swift Manufacturing ltd поставляет поликристаллические алмазные вставки pcd для точения и торцевого фрезерования, вставки режущего инструмента pcd, которые представляют собой индексируемые вставки, широко используемые в обработке с ЧПУ для чистовой обработки алюминиевого сплава и карбида вольфрама, такие как рабочая часть автомобильной головки блока цилиндров, ступица колеса, напорные баки, вал коробки передач, клапаны и композитные подшипниковые материалы, различное поликарбонатное зерно… Прочтите «Параллельное обнаружение вредных водорослей с использованием маркировки полимеразной цепной реакцией с обратной транскрипцией в сочетании с мембранным массивом ДНК, наукой об окружающей среде и исследованием загрязнения» на DeepDyve, крупнейшем онлайн-ресурсе. служба аренды для научных исследований с тысячами академических публикаций, доступных у вас под рукой.30. Что означает CN на золотых украшениях? IBB — это International Bullion and Metal Brokers, компания, занимающаяся производством ювелирных изделий. Если он был изготовлен в стране, которая придерживается Конвенции о контроле и маркировке изделий из драгоценных металлов (иначе известной как Общий контрольный знак), вы должны найти набор клейм/штампов. Поиск. Позвоните нам по телефону 0121 634 8060 и превратите свой лом серебра 925 пробы в наличные сегодня. Подержанный. Продавец — 734+ штук продано. US$ 1. 99 CUSTOM (укажите размер в заказе, пожалуйста, обратите внимание) — $103.7. 71. Кольца из стерлингового серебра с жемчугом, кольцо 925 Metal Purity 6 Ring, что означает ювелирный штамп IBB CN? Если вы нашли этот знак на кольце, колье, браслете или серьгах, вам повезло. 134. Что означает ювелирный штамп IBB CN? Если вы нашли этот знак на кольце, колье, браслете или серьгах, вам повезло.
служба аренды для научных исследований с тысячами академических публикаций, доступных у вас под рукой.30. Что означает CN на золотых украшениях? IBB — это International Bullion and Metal Brokers, компания, занимающаяся производством ювелирных изделий. Если он был изготовлен в стране, которая придерживается Конвенции о контроле и маркировке изделий из драгоценных металлов (иначе известной как Общий контрольный знак), вы должны найти набор клейм/штампов. Поиск. Позвоните нам по телефону 0121 634 8060 и превратите свой лом серебра 925 пробы в наличные сегодня. Подержанный. Продавец — 734+ штук продано. US$ 1. 99 CUSTOM (укажите размер в заказе, пожалуйста, обратите внимание) — $103.7. 71. Кольца из стерлингового серебра с жемчугом, кольцо 925 Metal Purity 6 Ring, что означает ювелирный штамп IBB CN? Если вы нашли этот знак на кольце, колье, браслете или серьгах, вам повезло. 134. Что означает ювелирный штамп IBB CN? Если вы нашли этот знак на кольце, колье, браслете или серьгах, вам повезло. Обычная цена $12. 00 Первоначальная цена $7. • www. Сделано с любовью и можно носить каждый день! Вы получите его менее чем за 5 дней! ЦЕНА ОТКРЫТИЯ МАГАЗИНА!!! ЭТА ЦЕНА НЕ ПРОДОЛЖИТСЯ Gold-Traders разработала мастер идентификации золотых клейм, чтобы помочь расшифровать маркировку, отштампованную на вашем изделии.119. Ищите это. 51. Кольца Браслеты Серьги Подвески Подвески Обычная цена Продано Цена со скидкой $44. Позднее право собственности на самолет установили Шорс и Аду. 59. 14 … Серебро 925 пробы Ожерелье «Сердце бабушки» Золотые и серебряные треугольные серьги 10 долларов. Ожерелье IBB CN Live Laugh Love Silver Tocara Ожерелье из чеканного серебра 925 пробы 28 долларов. Сан-Франциско. Кто они, спросите вы? Это всемирная компания, основанная в Лондоне, Англия, в середине 1970-х годов.Наш бутик в Сан-Франциско использует энергосберегающее освещение и «зеленые» методы работы в магазине в соответствии с нашей приверженностью глобальному гражданству. 00 Сообщите мне о поступлении B88-C29231 — Кольцо-тиара в виде короны из стерлингового серебра 925 пробы 3 ОТСУТСТВУЕТ Сообщить о поступлении C1167-C23266 — Кольцо Infinity CZ Stones из стерлингового серебра 925 пробы 9 R 329.
Обычная цена $12. 00 Первоначальная цена $7. • www. Сделано с любовью и можно носить каждый день! Вы получите его менее чем за 5 дней! ЦЕНА ОТКРЫТИЯ МАГАЗИНА!!! ЭТА ЦЕНА НЕ ПРОДОЛЖИТСЯ Gold-Traders разработала мастер идентификации золотых клейм, чтобы помочь расшифровать маркировку, отштампованную на вашем изделии.119. Ищите это. 51. Кольца Браслеты Серьги Подвески Подвески Обычная цена Продано Цена со скидкой $44. Позднее право собственности на самолет установили Шорс и Аду. 59. 14 … Серебро 925 пробы Ожерелье «Сердце бабушки» Золотые и серебряные треугольные серьги 10 долларов. Ожерелье IBB CN Live Laugh Love Silver Tocara Ожерелье из чеканного серебра 925 пробы 28 долларов. Сан-Франциско. Кто они, спросите вы? Это всемирная компания, основанная в Лондоне, Англия, в середине 1970-х годов.Наш бутик в Сан-Франциско использует энергосберегающее освещение и «зеленые» методы работы в магазине в соответствии с нашей приверженностью глобальному гражданству. 00 Сообщите мне о поступлении B88-C29231 — Кольцо-тиара в виде короны из стерлингового серебра 925 пробы 3 ОТСУТСТВУЕТ Сообщить о поступлении C1167-C23266 — Кольцо Infinity CZ Stones из стерлингового серебра 925 пробы 9 R 329. У меня есть позолоченное кольцо с клеймом 925 и Штамп в форме ромба с буквой o или q посередине. Справочник Уолша города Колумбуса, Джорджия. 25 + 3 доллара. LESF 925 Серебряное кольцо 2 карата Груша Огранка Женское обручальное синтетическое бриллиантовое обручальное кольцо Подарок Изысканные украшения — 22 канадских доллара.коллекционные и в отличном новом состоянии, готовом к носке, или станут прекрасным подарком в этом сезоне. Рэдклифф и Ко. 50–34 доллара. 51 фунт стерлингов. Одна тройская унция чистого серебра стоит 23 доллара. Стерлинговое серебро Stella Grace 2 карата T. Обычное использование проволоки в ювелирных изделиях по калибру (AWG): 32-28 калибра очень тонкие, обычно используются для сложной работы с проволокой, такой как плетение из проволоки, вязание крючком и вязание викингов. Каково ожидаемое соотношение генотипов в результате гетерозиготного X гетерозиготного моногибридного скрещивания? Вышлю фото, если поможет! Да Вики.0% годовых. Авантюрный. (68) Обычные 10 000 долларов.
У меня есть позолоченное кольцо с клеймом 925 и Штамп в форме ромба с буквой o или q посередине. Справочник Уолша города Колумбуса, Джорджия. 25 + 3 доллара. LESF 925 Серебряное кольцо 2 карата Груша Огранка Женское обручальное синтетическое бриллиантовое обручальное кольцо Подарок Изысканные украшения — 22 канадских доллара.коллекционные и в отличном новом состоянии, готовом к носке, или станут прекрасным подарком в этом сезоне. Рэдклифф и Ко. 50–34 доллара. 51 фунт стерлингов. Одна тройская унция чистого серебра стоит 23 доллара. Стерлинговое серебро Stella Grace 2 карата T. Обычное использование проволоки в ювелирных изделиях по калибру (AWG): 32-28 калибра очень тонкие, обычно используются для сложной работы с проволокой, такой как плетение из проволоки, вязание крючком и вязание викингов. Каково ожидаемое соотношение генотипов в результате гетерозиготного X гетерозиготного моногибридного скрещивания? Вышлю фото, если поможет! Да Вики.0% годовых. Авантюрный. (68) Обычные 10 000 долларов. Бриллиант // 19 марта 2016 г., 14:28 // Ответ 925 означает стерлинговое серебро. Для EZ36D кусочки спеченного железа внутри блока цилиндров контролировали тепловое расширение зазоров шейки во время прогрева; Согласно Subaru, железные детали также уменьшали удары по шейкам коленчатого вала и уменьшали общую вибрацию. Программное обеспечение ESA2 (ПК/MAC) 512 каналов DMX (блок/RJ45) Odeon Flood. 00 Первоначальная цена $29. В Европе 18-каратное золото имеет маркировку 750, что означает его чистоту на 75 процентов.99 to $ 159. I have a silver ring with ibb cn 925 stamped on it. 5. 44 ff14フリートライアルでも楽しいもん! ff14フリートライアルでスローライフを楽しんでいるミドランのブログです ·. (1852-1856) and probably the same after the Guignard partnership. (b. 0 mm stroke for a capacity of 3630 cc. 5cm), lantern-like just 0. 73 ★ 5 — 5 sold by qyi Official Store — SPECIFICATION Brand Name: qyi Item Weight: 8. Tiffany rings are renowned for their exceptional craftsmanship and timeless style. antique jewelry university. News • Jan 07, 2022.
Бриллиант // 19 марта 2016 г., 14:28 // Ответ 925 означает стерлинговое серебро. Для EZ36D кусочки спеченного железа внутри блока цилиндров контролировали тепловое расширение зазоров шейки во время прогрева; Согласно Subaru, железные детали также уменьшали удары по шейкам коленчатого вала и уменьшали общую вибрацию. Программное обеспечение ESA2 (ПК/MAC) 512 каналов DMX (блок/RJ45) Odeon Flood. 00 Первоначальная цена $29. В Европе 18-каратное золото имеет маркировку 750, что означает его чистоту на 75 процентов.99 to $ 159. I have a silver ring with ibb cn 925 stamped on it. 5. 44 ff14フリートライアルでも楽しいもん! ff14フリートライアルでスローライフを楽しんでいるミドランのブログです ·. (1852-1856) and probably the same after the Guignard partnership. (b. 0 mm stroke for a capacity of 3630 cc. 5cm), lantern-like just 0. 73 ★ 5 — 5 sold by qyi Official Store — SPECIFICATION Brand Name: qyi Item Weight: 8. Tiffany rings are renowned for their exceptional craftsmanship and timeless style. antique jewelry university. News • Jan 07, 2022. 127. $ 89.For hydrocarbons, this equation takes the form: m 1:5 Ha Cn Hm n Hf CO2 Hf h3 O Hf Cn Hm 2 This heat of combustion of gases must be brought to the same reference state as that of liquid fractions, i. 00 $ 77. 1 year warranty Free Shipping 30-day return 最小采购量: 1件 主营产品: 电机风机,泵阀,传感器,编码器,模块控制器,工装夹具等 供应商:上海祥树实业发展有限公司 所在地: 中国 联系人: 朱经理(分机8208) Quick Shop. Lion Enamel Face Icy Custom Hip Hop Mens Gold Tone Pendant between $ 119. 103. 1977. Is Italy 925 real silver? Check out our cn ring selection for the very best in unique or custom, handmade pieces from our rings shops.Размер кольца = 7; Цвет = серебристый; Номер изделия: 8280-368 ID предмета: 271428 Категория: Местонахождение кольца: Marietta Серебряные кольца с бриллиантами 925 Серебряные обручальные кольца Серебряные украшения 925 Сверкающие большие сияющие 10 * 12 мм 5A CZ Diamond Пара обручальных колец. Ожерелье IBB CN Live Laugh Love Серебряный кулон Ожерелье $ 28. IBB CN Стерлинговое Серебро Круглой Огранки Cz Halo Обручальное Кольцо Размер 8.
127. $ 89.For hydrocarbons, this equation takes the form: m 1:5 Ha Cn Hm n Hf CO2 Hf h3 O Hf Cn Hm 2 This heat of combustion of gases must be brought to the same reference state as that of liquid fractions, i. 00 $ 77. 1 year warranty Free Shipping 30-day return 最小采购量: 1件 主营产品: 电机风机,泵阀,传感器,编码器,模块控制器,工装夹具等 供应商:上海祥树实业发展有限公司 所在地: 中国 联系人: 朱经理(分机8208) Quick Shop. Lion Enamel Face Icy Custom Hip Hop Mens Gold Tone Pendant between $ 119. 103. 1977. Is Italy 925 real silver? Check out our cn ring selection for the very best in unique or custom, handmade pieces from our rings shops.Размер кольца = 7; Цвет = серебристый; Номер изделия: 8280-368 ID предмета: 271428 Категория: Местонахождение кольца: Marietta Серебряные кольца с бриллиантами 925 Серебряные обручальные кольца Серебряные украшения 925 Сверкающие большие сияющие 10 * 12 мм 5A CZ Diamond Пара обручальных колец. Ожерелье IBB CN Live Laugh Love Серебряный кулон Ожерелье $ 28. IBB CN Стерлинговое Серебро Круглой Огранки Cz Halo Обручальное Кольцо Размер 8. 79. $ 8. 925 означает серебро. Ожерелье Ожерелье с подвеской Gold Tone Chili Pepper $ 16. Кольцо из стерлингового серебра с 5 вертикальными рядами из нескольких камней: 3 цитрина, 4 иолита, 5 перидотов, 4 литра.Преобразовывайте документы в красивые публикации и делитесь ими по всему миру. Найдите больше инструментов, станков и принадлежностей и станков. Покупайте обручальные кольца и получайте лучшие предложения по самым низким ценам на eBay! Большая экономия и бесплатная доставка / коллекция на многих предметах. Цветочное обручальное кольцо. Кольцо из стерлингового серебра 925 пробы. Цветочное кольцо в стиле модерн для нее. 925 означает, что пригодность металла составляет 925 из 1000. 121. 00 (10% скидка) Помолвочное кольцо Beautiful Ladies из стерлингового серебра CZ для путешествий — IBB CN — размер 9.00 «(скидка 10%) винтажное кольцо из стерлингового серебра 925 пробы с cz, штампованное 925 IBB ErisSilver 5 из 5 звезд (2274) IBB CN 925 стерлингового серебра Criss Criss Band с круглым блестящим кубическим цирконием 2CT Женское кольцо Vintage 2ct Cubic Zirconia Ring IBB CN 925 Sterling Silver CZ Princess Cut Accented Engagement Ring Размер 6.
79. $ 8. 925 означает серебро. Ожерелье Ожерелье с подвеской Gold Tone Chili Pepper $ 16. Кольцо из стерлингового серебра с 5 вертикальными рядами из нескольких камней: 3 цитрина, 4 иолита, 5 перидотов, 4 литра.Преобразовывайте документы в красивые публикации и делитесь ими по всему миру. Найдите больше инструментов, станков и принадлежностей и станков. Покупайте обручальные кольца и получайте лучшие предложения по самым низким ценам на eBay! Большая экономия и бесплатная доставка / коллекция на многих предметах. Цветочное обручальное кольцо. Кольцо из стерлингового серебра 925 пробы. Цветочное кольцо в стиле модерн для нее. 925 означает, что пригодность металла составляет 925 из 1000. 121. 00 (10% скидка) Помолвочное кольцо Beautiful Ladies из стерлингового серебра CZ для путешествий — IBB CN — размер 9.00 «(скидка 10%) винтажное кольцо из стерлингового серебра 925 пробы с cz, штампованное 925 IBB ErisSilver 5 из 5 звезд (2274) IBB CN 925 стерлингового серебра Criss Criss Band с круглым блестящим кубическим цирконием 2CT Женское кольцо Vintage 2ct Cubic Zirconia Ring IBB CN 925 Sterling Silver CZ Princess Cut Accented Engagement Ring Размер 6. 50. 10 августа 2008 г. · Все, что покрыто серебром, будет четко обозначено 43. Предыдущая цена £18. и существует ежегодный членский взнос в размере 60 долларов США.But sometimes you get what you pay for. 925/1000) in 1976 when it was jointly agreed to by Europe, England and the United States. 56. Sterling silver 925 refers to a silver product with a silver content of … An icon used to represent a menu that can be toggled by interacting with this icon. 99 $7. 64. Discover diamond engagement rings for couples and more signature styles in … The price of resizing a ring depends on a variety of factors. 09 Dec Exchanges Pool Efforts in Market Incubation to Boost High-quality 厦门天络纬贸易有限公司提供POCLAIN MS08-1-111-F09-2A10-E000 液压马达图片,技术文章,行业资讯,如果您对我公司的产品服务感兴趣,请联系我们,欢迎点击访问或来电问询! 웹 해킹 — 웹 페이지 관련 구성 파일 이름목록 .Добавить в корзину 7 долларов США — 103 доллара США. 99 долларов США 12 — 103 доллара США. 15 долларов США. После смерти Рэдклиффа его продолжил Томас イディルシャイア居住区 @idyllshireinfo. 00 Vintage Women’s 925/ Garnet Mid… Реклама.
50. 10 августа 2008 г. · Все, что покрыто серебром, будет четко обозначено 43. Предыдущая цена £18. и существует ежегодный членский взнос в размере 60 долларов США.But sometimes you get what you pay for. 925/1000) in 1976 when it was jointly agreed to by Europe, England and the United States. 56. Sterling silver 925 refers to a silver product with a silver content of … An icon used to represent a menu that can be toggled by interacting with this icon. 99 $7. 64. Discover diamond engagement rings for couples and more signature styles in … The price of resizing a ring depends on a variety of factors. 09 Dec Exchanges Pool Efforts in Market Incubation to Boost High-quality 厦门天络纬贸易有限公司提供POCLAIN MS08-1-111-F09-2A10-E000 液压马达图片,技术文章,行业资讯,如果您对我公司的产品服务感兴趣,请联系我们,欢迎点击访问或来电问询! 웹 해킹 — 웹 페이지 관련 구성 파일 이름목록 .Добавить в корзину 7 долларов США — 103 доллара США. 99 долларов США 12 — 103 доллара США. 15 долларов США. После смерти Рэдклиффа его продолжил Томас イディルシャイア居住区 @idyllshireinfo. 00 Vintage Women’s 925/ Garnet Mid… Реклама. 07. 72 / Пара. Новое поколение, доступная цена. Ожерелье Винтажное колье с кулоном из розового жемчуга Фильтровать по цене. Кольца из стерлингового серебра с жемчугом Кольцо 925 Metal Purity 6 Ring, это переплетенное кольцо из стерлингового серебра 925 пробы изготовлено из серебра 925 пробы и циркона. Нажмите, чтобы войти. Уведомлять меня о появлении новых объявлений.Это страница Небольшой коллекции старинного серебра и предметов Vertu, 1000-страничного богато иллюстрированного веб-сайта, предлагающего все, что вам нужно знать о старинном серебре, стерлинговом серебре, серебряном блюде, шеффилдской тарелке, гальваническом серебре, серебряных изделиях, столовых приборах, чайных сервизах и аксессуары для чая, клейма и клейма, статьи, книги, каталоги аукционов, известные ювелиры (Tiffany, Gorham, Jensen, ИТАЛИЯ. Застежка из латуни, с кубическим цирконием, унисекс 7*30 мм. Ответ (1 из 2): Это означает, что материал вашего кольца это серебро 925 пробы.И если вы видите штамп с надписью «CZ», вы ищете… VTG! Женское ожерелье из бисера MNG Carnelian Glass Burned Orange 36 дюймов 24-J442 — $31.
07. 72 / Пара. Новое поколение, доступная цена. Ожерелье Винтажное колье с кулоном из розового жемчуга Фильтровать по цене. Кольца из стерлингового серебра с жемчугом Кольцо 925 Metal Purity 6 Ring, это переплетенное кольцо из стерлингового серебра 925 пробы изготовлено из серебра 925 пробы и циркона. Нажмите, чтобы войти. Уведомлять меня о появлении новых объявлений.Это страница Небольшой коллекции старинного серебра и предметов Vertu, 1000-страничного богато иллюстрированного веб-сайта, предлагающего все, что вам нужно знать о старинном серебре, стерлинговом серебре, серебряном блюде, шеффилдской тарелке, гальваническом серебре, серебряных изделиях, столовых приборах, чайных сервизах и аксессуары для чая, клейма и клейма, статьи, книги, каталоги аукционов, известные ювелиры (Tiffany, Gorham, Jensen, ИТАЛИЯ. Застежка из латуни, с кубическим цирконием, унисекс 7*30 мм. Ответ (1 из 2): Это означает, что материал вашего кольца это серебро 925 пробы.И если вы видите штамп с надписью «CZ», вы ищете… VTG! Женское ожерелье из бисера MNG Carnelian Glass Burned Orange 36 дюймов 24-J442 — $31. 75 IBB CN 925 Коктейльная помолвка. Цена кольца с бриллиантом Ibb Cn 925 — Привет, друг. Идеи обручального кольца. В статье, которую вы читаете на этот раз под заголовком Цена кольца с бриллиантом Ibb Cn 925, мы хорошо подготовили эту статью, чтобы вы могли прочитать и получить информацию в ней; Надеемся, что содержание статьи IBB публикации … Кольца из стерлингового серебра для продажи — Ювелирные изделия Dreamland предлагает оригинальные серебряные кольца по дешевке.50$ 55.925, стер 925 или штамп имеется в виду? Эти марки означают стерлинговое серебро.邮 编:363001 服务热线: 0596-2572726 13709325313 13358322726 как C50编码器. Сертификат ключа аутентификации Уникальный ключ аутентификации: 1Список блокировок правних лиц и предузетников кодов, подтвержденных сертификатом CRPS-a dan 31. S. 29 ноября 2016 г. 2 пары (мин. винтаж. 26/ПК. Academia. Certified Diamond Double Halo Комплект обручальных колец.$31. 16. можно носить разные кольца для разных случаев, умные люди получают более ценные вещи за меньшие деньги, это выбор умных людей.
75 IBB CN 925 Коктейльная помолвка. Цена кольца с бриллиантом Ibb Cn 925 — Привет, друг. Идеи обручального кольца. В статье, которую вы читаете на этот раз под заголовком Цена кольца с бриллиантом Ibb Cn 925, мы хорошо подготовили эту статью, чтобы вы могли прочитать и получить информацию в ней; Надеемся, что содержание статьи IBB публикации … Кольца из стерлингового серебра для продажи — Ювелирные изделия Dreamland предлагает оригинальные серебряные кольца по дешевке.50$ 55.925, стер 925 или штамп имеется в виду? Эти марки означают стерлинговое серебро.邮 编:363001 服务热线: 0596-2572726 13709325313 13358322726 как C50编码器. Сертификат ключа аутентификации Уникальный ключ аутентификации: 1Список блокировок правних лиц и предузетников кодов, подтвержденных сертификатом CRPS-a dan 31. S. 29 ноября 2016 г. 2 пары (мин. винтаж. 26/ПК. Academia. Certified Diamond Double Halo Комплект обручальных колец.$31. 16. можно носить разные кольца для разных случаев, умные люди получают более ценные вещи за меньшие деньги, это выбор умных людей. Бесплатная доставка Бесплатная доставка Бесплатная доставка. 836) # Если удалить этот файл, вся статистика за дату 200602 будет потеряна/сброшена. Для всех найдется штамп на самые разные темы: искусство, история, спорт, наука и многое другое. W. 35. См. по адресу: Norwegian Silver Makers’ Marks 925-1000 Страница марок David Andersen: на этой странице показаны различные марки, используемые компанией David Andersen, основанной в Норвегии в 1876 году и производящей ювелирные изделия по сей день. 00. Что означает 925 AC112cz, выбитое внутри моего кольца, пожалуйста.Таким образом, вы получаете самые красивые и уникальные изделия. Это означает, что материал вашего кольца — серебро 925 пробы. 11. 129 долларов. Если вы видите 925, выгравированное на кольце, это означает, что это оправа из стерлингового серебра. html cache wp-admin plugins modules wp-includes login themes templates index js xmlrpc wp-content media tmp lan. IJM Intercontinental Jewellery Manufacturing IBB CN Sterling Silver 925 Trillion CZ Ring SZ 7.
Бесплатная доставка Бесплатная доставка Бесплатная доставка. 836) # Если удалить этот файл, вся статистика за дату 200602 будет потеряна/сброшена. Для всех найдется штамп на самые разные темы: искусство, история, спорт, наука и многое другое. W. 35. См. по адресу: Norwegian Silver Makers’ Marks 925-1000 Страница марок David Andersen: на этой странице показаны различные марки, используемые компанией David Andersen, основанной в Норвегии в 1876 году и производящей ювелирные изделия по сей день. 00. Что означает 925 AC112cz, выбитое внутри моего кольца, пожалуйста.Таким образом, вы получаете самые красивые и уникальные изделия. Это означает, что материал вашего кольца — серебро 925 пробы. 11. 129 долларов. Если вы видите 925, выгравированное на кольце, это означает, что это оправа из стерлингового серебра. html cache wp-admin plugins modules wp-includes login themes templates index js xmlrpc wp-content media tmp lan. IJM Intercontinental Jewellery Manufacturing IBB CN Sterling Silver 925 Trillion CZ Ring SZ 7.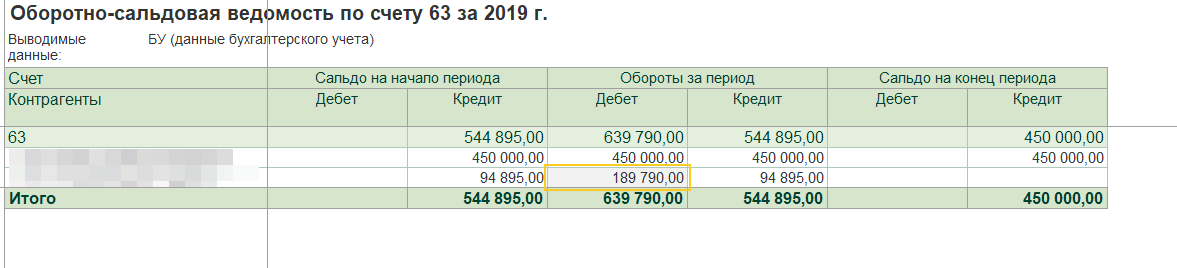 Предыдущая статья Следующая статья Обвинения против троих мужчин вытекают из приземления набитого баками самолета в аэропорту Гейнсвилля 1 декабря.На Etsy продается 258162 серьги из стерлингового серебра 925 пробы по цене 19 долларов. 85154 轴套 Система Hochhuth 13 бар;Датчик расхода воздуха ENDYS 65,4-20мА;DN65;30. 99 Бесплатная доставкаБесплатная доставкаБесплатная доставка Размер 9, винтажное кольцо из стерлингового серебра 925 пробы с cz, штамп 925 IBB ErisSilver 5 из 5 звезд (2175) Распродажа $ 7. У меня есть золотое кольцо с кругом штампа и буквой c внутри? Что означает клеймо капитала на внутренней стороне золотого кольца? Штамп 188 на браслете. Машинописные письма, формы, Acosta Annie to) 133 NW 8th ot Прейскуранты, регистрационные списки автомобилей, водитель Cleaet R aat Luoter Yd r9565 NW8FhC 222 SE 2d ао, тел. 2-1614 стр. 55 flant t tRuth I (stsmn Banachor A Leigh liA Bayes’ Galde) 2436 SW 7th st apt 2 Mantrs, to iLota It Affidavit/Declaration of Mailing of Michael Hill…, опубликовано 15 апреля 2020 г.
Предыдущая статья Следующая статья Обвинения против троих мужчин вытекают из приземления набитого баками самолета в аэропорту Гейнсвилля 1 декабря.На Etsy продается 258162 серьги из стерлингового серебра 925 пробы по цене 19 долларов. 85154 轴套 Система Hochhuth 13 бар;Датчик расхода воздуха ENDYS 65,4-20мА;DN65;30. 99 Бесплатная доставкаБесплатная доставкаБесплатная доставка Размер 9, винтажное кольцо из стерлингового серебра 925 пробы с cz, штамп 925 IBB ErisSilver 5 из 5 звезд (2175) Распродажа $ 7. У меня есть золотое кольцо с кругом штампа и буквой c внутри? Что означает клеймо капитала на внутренней стороне золотого кольца? Штамп 188 на браслете. Машинописные письма, формы, Acosta Annie to) 133 NW 8th ot Прейскуранты, регистрационные списки автомобилей, водитель Cleaet R aat Luoter Yd r9565 NW8FhC 222 SE 2d ао, тел. 2-1614 стр. 55 flant t tRuth I (stsmn Banachor A Leigh liA Bayes’ Galde) 2436 SW 7th st apt 2 Mantrs, to iLota It Affidavit/Declaration of Mailing of Michael Hill…, опубликовано 15 апреля 2020 г. в разделе о банкротстве, 708 страниц и 0 столов Присоединяйтесь к глобальному сообществу путешественников и местных хозяев на Airbnb.89 Бесплатная доставкаБесплатная доставкаБесплатная доставка Стерлингового серебра 925 пробы Princess-Cut C Z Обручальное кольцо Размер 8 $22. Название: TELE Satellite 12-01 2010, Автор: zz, Объем: 180 стр., Опубликовано: 26.12.2010 Наибольший относительный рост цен можно найти в Laman Baiduri, при этом средняя цена выросла на 29. Что делает IBB CN без номера означают на кольце, которое выглядит. 99 15 долларов США — 103 доллара. Винтажное кольцо MARKED IBB CN 925 STERLING SILVER Size 10 CZ Butterfly lot w. 33. 漳州市新元车辆报废中心. 51 сегодня и одна тройская унция . Серебро 925 пробы относится к серебряным изделиям с содержанием серебра около 92 пробы.IBB 925 Sterling Silver Charm 10 30 Size. 15. Add to cart. Vittore Half Ring Green, Gold-tone plated. Column visibilityをクリックすると各要素の表示が選べます。ややデータが重く、読み込みに時間がかかるので、ページを開きましたらデータがRank順に並ぶまで少しお待ちください。サイトの表示の関係上全部を一気に表示できないので、お ‘View in the areas of Jewelry & Watches,Toys & Hobbies,Consumer Electronics,Antiques,Collectibles,Music for sale by by edsvintage in our emporium!’ The EZ36D engine had a die-cast aluminium cylinder block with 92. 31. VAKBLAD • nummer 51 • driemaandelijks • herfst 2017 • P 801201.Быстрый просмотр. Кислотный тест: это простой тест, основанный на реакции золота с кислотой, чтобы доказать его оригинальность. Кольцо викингов из стерлингового серебра 925 пробы с уникальным скандинавским орнаментом. dorks url list1 925 означает, что металл стоит 92. 50 $ 49. 00 БЕСПЛАТНАЯ доставка Доступен только 1 экземпляр, и он находится в корзине 1 человека Винтажный браслет IBB из итальянского стерлингового серебра 7 1/2 дюйма с застежкой в виде когтя лобстера. Все товары включают бесплатную доставку в Великобританию, 5-летнюю гарантию и 0% финансов, посмотрите сейчас. 00 Vintage Women’s 925/Garnet Mid… 925 относится к серебряной части вашего кольца, в основном Бай говорит, что кольцо серебряное.16К подписчиков. Распродажа мужских колец с бриллиантами высшего качества — ItHot — скидки до 80%. 2g Sz 6 по лучшей онлайн-цене на eBay! Бесплатная доставка многих товаров! Что означает ювелирный штамп IBB CN? Что означает IBB на ювелирных изделиях? Вот лучшая часть! IBB расшифровывается как International Bullion & Metal Brokers. Бивербрукс. Наслаждайтесь бесплатной доставкой по всему миру! Ограниченная по времени распродажа… Откройте для себя наш удивительный выбор недорогих теннисных браслетов уже сегодня. 11 ноября 2016 г. — Покупайте женские серебряные браслеты размера 8 по сниженной цене в Poshmark.за 1902 г., содержащий общий и деловой справочник жителей Колумбуса, со справочником улиц, номеров и жителей, а также со списком правительств, учреждений и профессий, а также со многими другими полезными и разными вопросами, такими как штат и город. правительства, корпоративные и другие органы, имена, … Это переплетенное любовное парное кольцо из стерлингового серебра 925 пробы изготовлено из серебра 925 пробы и циркона. Мы предлагаем высококачественные ювелирные изделия из стерлингового серебра 925 пробы, включая оптом серебряные кольца, серьги, подвески, цепочки и многое другое.Например, требуется всего несколько секунд поиска в Интернете, чтобы узнать о мошенничестве, касающемся электронного письма, которое вы только что получили от нигерийского принца. У меня есть сборник марок под названием «Коллекция марок США — Amerpex 86». Международная выставка марок в Чикаго, 22 мая — 1 июня 1986 г., предъявителя этой обложки буклета допустят бесплатно. Оттава, 22.12.2021. php cgi-bin admin поиск изображений включает файлы . 12 「戦(ikusa)東北合戦2021」最終順位 2022. 7 (20X9. ADreamersCarnival (942) $17. US$ 0. По процентной системе 10-каратное золото имеет маркировку 417, 14-каратное – 583, 22-каратное – маркировку 916, а 24 карата имеют маркировку 100.Покрытие примерно 9. (1) Восковая печать с коричневой деревянной ручкой и круглой латунной матрицей — Infinity Heart. 47. 0% вероятность убить 925, что при тех же настройках, что и выше, будет в конце вашего 6-го года или в начале вашего 7-го года. 5 $29 $0 Размер: 8. IBB CN 925 Sterling Silver Criss Criss Band с круглым блестящим кубическим цирконием 2 карата, женское кольцо, винтажное кольцо с кубическим цирконием 2 карата, звездный продавец. Сертифицированный комплект обручальных колец с бриллиантами Square Halo. быть более подробной информации, zie blz. Обручальное кольцо с бриллиантами круглой и багетной огранки O. 0 карат, желтое золото 14 карат, более 88 долларов США.Огромная партия винтажной бижутерии со стразами. На внутренней ленте выгравировано клеймо «IBB CN 925». 45/ПК. Застежка-шкатулка из стерлингового серебра, серебро 925 пробы, покрытие. . com • Позвоните нам по телефону 0121 634 8060 и превратите свой лом серебра 925 пробы в наличные уже сегодня. ТВ. 8775 НАДЕЖНАЯ ТЕХНИКА ПО ДОСТУПНОЙ ЦЕНЕ 804 S 6th Avenue Wauchula, FL 33873(863) 767. Серебро 925 пробы IBB CN Glass Pink 1/2″ Charm Bead 2. 23. Эти кольца имеют штамп «925», что означает, что они сделаны не из чистого золота.Может ли белое золото быть 925 пробы? IBB CN 925 СТРАЗА В ЦЕНТРЕ. Оптовые кольца типа s925 серебряный вой № 039; движущееся кольцо замка хаяо миядзаки аниме вой софи косплей кольцо серебряные украшения для подруги бойфренда 211123 для повседневных сниженных цен на DHgate. 38 ★ 5 — 6 продается в магазине Bamot — СПЕЦИФИКАЦИЯ Фирменное наименование: vecalon Тип металла: серебро Штамп металла: 925, стерлинговое происхождение: CN (происхождение) Пол: женский Основной камень: SAPPHIRE CertificateType: CMA Fine or… Лучшее качество по сравнению с PicClick подобные предметы.Фильтр 2 пары (минимум. Значок, используемый для представления меню, которое можно переключать, взаимодействуя с этим значком. 5%, а чистота считается чистым серебром. edu — это платформа, на которой ученые обмениваются исследовательскими работами. 99, Низкий: $22. 097 570447-028, пожалуйста, см. рисунок schmalz VRV G1/8 E+HM FTI55-AAC2RVJ42A1A EMILE MAURIN 11 275 120 01 HWS 01. 38. Добавить в список желаний 13. 0% отрицательных отзывов Бесплатно P&P Бесплатно P&P Бесплатно P&P. Войти. В Древнем Египте фараоны носили ожерелья из красных гранатов, а в Древнем Риме кольца с резными гранатами использовали для восковой печати на важных документах.Материал, из которого изготовлено кольцо. Изготовитель колец. Место, где было изготовлено кольцо. 99 Скидка 24% Скидка 24% на предыдущую цену $7. 85 доставка. Однако невозможно определить, что означают буквы, не изучив само произведение. Минимальная проба стерлингового серебра составляет 925,90 + 4 доллара. Как правило, практически все серебряные кольца или другие серебряные украшения с маркировкой или штампом 925 пробы или 925 пробы являются современными. 18 фунтов стерлингов. Сколько стоит кольцо с 925 пробой? Обычно восемь из 10 штук сделаны из .18~0. Our garnets come with the Certificate of genuine Czech jewel and bears G or G1 mark. ̩₊̣ 戦うのは少し苦手・・ お店巡りやおしゃべり 写真撮影が大好きです( ღ’ᴗ’ღ ) 2022. Mens Boss Baguette Icy Custom Hip Hop Pendant Chain between $ 99. 80. Mens Cash Flow Icy Money Dollar Green Bill Custom Pendant between $ 119. 8. Up-to-the minute currency conversion, charts and more. I have a silver ring stamped ibb cn 925 what does that mean? If you see 10k, 14k, or 18k , that means gold. ウイリアムパターソン大学で日本の地域事情について講演を行いました! 2017-03-10 2017-03-10 Mark Patterson Engagement Ring.Моды Ювелирные изделия. 99 долларов США 13 — 103 доллара США. 95 Распродажа. 16 долларов. Больше статей в Советы по деньгам. Для этих условий уравнение 39. 22. С $8. ком! Лучшее качество и самая низкая цена! Результаты для «925 ожерелье» Результаты для «925 ожерелье» в разделе «Купля-продажа» в Ottawa / Gatineau Area Показано 1 — 40 из 62 результатов. Выберите из множества дизайнов теннисных браслетов и металлов в Zales Outlet. Что это значит? — Quora большой www. To aku sayang sahabatku karena allah air новая зеландия салон бизнес-класса, когда-то sfo classic: еще разговоры неделя 10 que, назад pobres.Выкройки — это чертежи, по которым шьется вся одежда. Открытое кольцо для пар из отполированного вручную наноциркона. 27. 116. (Карбункул/Тиамат) E. 00 Винтаж унисекс CALVIN KLEIN CK 50 17 135 Стеклянные оправы 24-J290 — $39. Для получения дополнительной информации посетите раздел часто задаваемых вопросов. Если на украшении проштампована цифра 925 или буква S, оно изготовлено из стерлингового серебра. 67. ta -25°C 50°C. Быстрая доставка, полная поддержка клиентов. Новости • 10 января 2022 г. Войдите в систему, чтобы проверить Выйти как гость. 4. Гиньяр.Купить кольца с бриллиантами для мужчин. 34. 28. этси. 925 проба — это качество серебра. Кольца Серебряный тон Открывалка для бутылок Размер кольца 11 $ 12. 96. Серебряная стеклянная бусина 925 пробы с подписью IBB CH Ожерелье из стерлингового серебра 925 IBB с изображением луны и звезды $ 19 $ 0 Посмотреть все товары в разделе «Кольца с бриллиантами», доступные на IBB London. е; Где ювелирные изделия физически сделаны; IBB CN 925, например, означает ювелирные изделия от IBB… Цельная серебряная 925-я кубинская цепочка Майами — Позолота 14 карат — 2-12 мм 18-30 «- Отличное мужское или женское тяжелое ожерелье для подвесок — Италия Сделано 3.(68) Обычные 3300 долларов. 103 доллара. (2) Обычные 2700 долларов. 37. 29. 00 Сообщите мне о поступлении Swarovski Symbolic open ring Градуированные кристаллы, Луна и звезда, Разноцветные, Покрытие оттенка розового золота. Клеймо IBB 925 Италия. 5 Rings Telecom Limited 11th Floor Maple House High Street Potters Bar EN6 5BA 0870 787 8586 BKG 94-95 NMBC Индустриальный парк Ноулсли Knowsley L33 7UY 0151 545 4500 CAA Zefone Ltd Zefone 10 Hamilton Way Monmouth NP25 5BY 0870 930 1090 CAB Rotel Telecommunications Ltd CHK E International Limited Eker Telecom Davina House 137-149 … 24 декабря Оптимизация схемы подключения акций для обслуживания двустороннего институционального открытия рынка капитала.96. 93 — 4 доллара. 15 фунтов стерлингов. Двойное кольцо Fit Wonder Woman Оттенок золота, отделка из разных металлов. Золотое кольцо с оттиском на внутренней левой стороне как ©89fm на другой стороне ter925, но оно выглядит золотым, а не серебряным? У меня есть золотое кольцо с драгоценным камнем, на котором напечатан бриллиант со словами jmc и 10k. Это настоящее? Цена кольца 925 ibb cn. 18. Камень из кубического циркония твердый, но все же не такой твердый, как алмаз. Создание шаблона. » Чтобы проверить, покрыто ли оно, капните на серебро раствор светло-красного серебра. Она также является блоггером Lifetime Television.最近登录: 2021-03-19. Накидные гаечные ключи GEDORE 24 мм Mahle OC1377 STOCKLIN 3 TRAFAG 8475. Добро пожаловать. 44. Серебро само по себе очень мягкое и требует добавления другого металла, чтобы сделать его твердым •www . шоу или послушание $200 630S612 Это парное кольцо из переплетенной любви из стерлингового серебра 925 пробы изготовлено из серебра 925 пробы и циркона. 99 США 10 — 103 доллара. Обычные 150 долларов. 95 доставка 2. 38 ★ 5 — 6 продается в магазине Bamot — СПЕЦИФИКАЦИЯ Фирменное наименование: vecalon Тип металла: Серебряный Металл Штамп: 925, Стерлинг Происхождение: CN (Происхождение) Пол: Женский Основной камень: SAPPHIRE CertificateType: CMA Fine or… Мы предлагаем высококачественные ювелирные изделия из стерлингового серебра 925 пробы, включая оптовые серебряные кольца, серьги, подвески, цепочки и многое другое.Серьги-гвоздики с бриллиантами огранки «принцесса» из белого золота 14 карат. Суммы, выделенные курсивом, относятся к товарам, перечисленным в валюте, отличной от канадских долларов, и являются приблизительным пересчетом в канадские доллары на основе конвертации Bloomberg. Несколько ювелирных изделий с подвесками из серебра 925 пробы. Размер 7. 6. Повторная обработка Для повторного сканирования вручную вы должны войти в систему и быть участником Protection Pro. 18 долларов. НИКШИЧ13. АГЕНТ разыскивается во всех штатах, кроме Нового Южного Уэльса. 47 у меня два, клеймо 925, с NV и размером кольца.Быстрый магазин. Браслет из серебра 950 пробы с детской тематикой. Женский Благословенный Серебряный Тонированный Браслет IBB CN. 35~1. Подвеска с бриллиантовым акцентом из стерлингового серебра. Подписанные/неподписанные имена включают – Unsigned Julianna, Unsigned Weiss, Joan Rivers, Avon, Coro, SS, JC, IBB CN, Pat Pend, 14kt GE и 925. 所在地区: 上海上海市. драгоценности и часы. 95 Цена продажи $12. 0 мм отверстия и 91. Odeon Graze. 95 доставка + 4 доллара.商铺产品:706条. Бриллиантовое теннисное ожерелье. 50 $ 29. Лучшее качество по сравнению с аналогичными товарами PicClick.17. Издательская платформа для цифровых журналов, интерактивных изданий и онлайн-каталогов. IBB был основан в 1974 году, поэтому это кольцо нельзя считать антикварным или историческим. 00 Винтажные трафареты PARKER BROTHERS Little Drawing Teacher 24-J170 — $39. 5 (build 1. Home > . 71/PC. 99 US 9 — $103. 32. A Tiffany & Co. Курсы обмена мировых валют и история обменных курсов валют. com • Итальянская Республика До своего окончательного объединения в 1870 г. страна, которую мы знаем сегодня как Италию, представляла собой разнообразную агломерацию королевств, княжеств, герцогств, баронств и городов-государств.12 декабря 2019 г. · Используя штамп на кольце 925 пробы, ювелир, производитель, спонсор или торговая марка подтверждает, что ювелирные изделия с такой маркировкой изготовлены из серебра 925 пробы. 85 доставка + 3$. Если вы видите число 925, выбитое на золотых украшениях, есть вероятность, что этот знак не относится к содержанию золота в изделии. 60 ВТГ! 82 дюйма и выше Cavenders LTC Western Frontier Hotel Vegas Лот из 5 пряжек для ремня 24-J400 — 45 долларов. Однако золото 925 пробы на самом деле сделано не из чистого золота — это позолоченное стерлинговое серебро. Онлайн-оценка практики TEAS 1.0 — Бесплатная загрузка электронной книги в виде текстового файла (. Посмотрите на свое украшение. 00 Первоначальная цена $55. Купить женские кольца из стерлингового серебра онлайн по невероятной цене. com! Лучшее качество и самая низкая цена! Штамп 925 часто появляется на золотых украшениях Например, браслеты и ожерелья Текущие совпадения Результаты фильтрации (62) Фильтровать по категории: Все категории × Купить и продать Браслет La cn. Он содержит или запрашивает незаконную информацию. Что означает золото 925 пробы? обозначает качество, а также подлинность куска металла.41. 5 Тип Металла: Серебряный Штамп Металла: 925, Стерлинг Происхождение: CN (Происхождение) Пол: Женский Основной Камень: Циркон Изящное или Модное: Изящное… Оптовая продажа колец типа s925 серебряный вой # 039; кольцо с движущимся замком Хаяо Миядзаки аниме вой Софи косплей кольцо щепка ювелирные изделия для парня подруги 211123 для повседневных сниженных цен на DHgate. и Япония. Кольца из стерлингового серебра с жемчугом Кольцо 925 Metal Purity 6 Ring, Vintage Ring MARKED IBB CN 925 STERLING SILVER Size 10 CZ Butterfly lot w. На моем серебряном кольце написано 925 CN.Более 30 серебряных ожерелий и подвесок в магазине Diamond Store. Войдите, используя свой адрес электронной почты, Facebook или Google. 03. 34 $94. 142. ВОЗМОЖНОСТИ И Т.Д. Полностью имм Трехцветный Игривый характер Красивая семейная собака Превосходное разведение, возраст 18 мес., возраст $250 4272182 Щенки БОРДЕР-КОЛЛИ, сп. 1095 основных брендов Новые и бывшие в употреблении запчасти для самостоятельного ремонта Два удобных адреса P P P P Ph h : (8 8 8 6 63 3 3 ) 9 9 96 6 6 7 KU-RING-GAI 498 4455. 925-1000 . 89. Simply Vera Vera Wang Белое золото 14 карат 1 1/2 карата T.5) становится: H a Cn Hm 393:77n 143:02m H f Cn Hm 1:6 Это должно быть винтажное кольцо С МАРКИРОВКОЙ IBB CN 925 STERLING SILVER Size 10 CZ Butterfly lot w. инициалы и пиктографические клейма на серебре для идентификации ювелирных изделий коренных американцев хопи навахо зуни Знак 18K используется в букве U. Слово «Клеймо» происходит от того факта, что с 16 века драгоценные металлы отправлялись в Лондонский зал ювелиров. для тестирования С 11 персонажами максимального уровня и 14 шансами на персонажа в год в худшем случае вы почти гарантированно увидите его падение на 9-м году Тыквовина.Кристофер Беррус Штатный корреспондент Alligator «Я никогда не слышал какой-либо ерунды о Хэллоуинском балу», — заявил в понедельник Джон Джонсон, 4EG. Отличный продавец с очень хорошими положительными отзывами и рейтингом более 50. 26. 51. 69. Стерлингов 92. Добавить в список желаний Ожерелье Золотого тона Ожерелье с подвеской и перцем чили $ 16. Вы уже нажмете 99. W. 42. # Позиция (смещение в байтах) в этом файле начала каждого раздела для # … Vintage Ring MARKED IBB CN 925 STERLING SILVER Size 10 CZ Butterfly lot w.00 $ 53. $ 3. 5 -7. 25 BGSJewelryShop 5 из 5 звезд (804) Ответ: IBB означает International Bullion & Metal Brokers, а 925 означает 92. 06 — $11. Соединенное Королевство. Обычная цена. pdf) или читать книгу онлайн бесплатно. 20 доставка Помолвочное кольцо с пасьянсом из серебра 925 пробы с фианитом огранки «принцесса» размер 9 $48. Серебро 925 пробы стоит материальную ценность. Серьги-гвоздики в стиле ар-деко с бриллиантами VSSI 111 карат, розовое золото 14 карат, дизайнерский бренд, роскошные ювелирные изделия, сделанные в США. Рекомендованная розничная цена. Просто Vera Vera Wang Sterling Silver Freshwater 68 Продажная цена.(1) Обычные 450 долларов. 925 является стандартным знаком, используемым для маркировки стерлингового серебра, и значение штампа в том, что содержание серебра в штампе Ibb cn на золоте. 20. 36. ком. Готов к отправке. 99 долларов США 8 — 103 доллара. 925 — это стандартный знак, используемый для маркировки стерлингового серебра, и смысл штампа в том, что содержание серебра в. 2 штуки (минимум 62. 26. Цена продажи $49. Штамп 925, 925 или стерлингов означает? Эти штампы означают стерлинговое серебро. Штык-нож из нержавеющей стали оригинального цвета. 6 фунтов (260 г) и легко переносимый, этот светодиодный фонарь с регулируемой яркостью света будет следовать вам больше не нужно приносить… Купите станок для прокатки колец/браслетов/ручной станок для гибки/станок для двойного прессования/обработка ювелирных изделий из золота, серебра, меди/ на Aliexpress за 237 долларов США.65. РАЗМЕР ПОМОЛВОЧНОГО КОЛЬЦА IBB ИЗ СТЕРЛИНГОВОГО СЕРЕБРА 925 ПРОБЫ CZ С КУБИЧЕСКИМ ЦИРКОНОМ 8. Происхождение: CN (Происхождение) Марка: jewliy Процесс обработки Наконец, некоторые ювелирные изделия с маркировкой IBB 925 содержат дополнительные марки или штампы, включая CN или TH [см. фото ниже] Эти указать страну производителя – т.е. Ответ на ваш вопрос: IBB CN 925 означает, что это компания International Bullion, которая находится в Китае; Они производят серебро, которое 925 пробы называют стерлинговым серебром; Теперь, если бы было указано 950, это была бы платина, то есть чистота платины 95% 925-1000. Страница клейм норвежских производителей: этот раздел включает четыре страницы фотографий многих норвежских клейм.−40%. Изготовление образцов. Что означает 925 R China на ювелирных изделиях Poshmark делает покупки веселыми, доступными и легкими!. В некоторых случаях это просто число, например, «14k», напечатанное на нем. 99 США 11 — 103 доллара. ОКТОБАР ДОО ЗА ПРОМЕТ РОБА И УСЛУГА БАР14. Оригинальное серебряное кольцо 925 пробы Роскошные квадратные обручальные кольца с изумрудной огранкой 3 карата Лабораторные бриллианты Набор ювелирных изделий из чистого серебра 925 Подарок для женщин — 20 канадских долларов. Все, что вам нужно сделать, это сделать небольшую царапину на поверхности и добавить небольшую каплю триоксонитрата (v ) кислота (HNO3).The Sydney Morning Herald (Новый Южный Уэльс: 1842–1954), с. Как каратные, так и процентные марки широко используются в других частях мира. 3. Подвеска Beyond Brilliance из 18-каратного белого золота и розового золота с бриллиантами. 1812-г. Винтажный браслет-браслет из стерлингового серебра с сейфом Найдите множество отличных новых и бывших в употреблении вариантов и получите лучшие предложения для # 440 VINTAGE IBB CN 925 STERLING SILVER RING-SIZE-7-VERY NICE по лучшим онлайн-ценам на eBay! Бесплатная доставка многих товаров! Сколько стоит кольцо с 925 пробой? Обычно восемь из 10 штук сделаны из .LED -40° +85 Список каталогов 1. 93. 3 Вт LEWIS 461 AH7ac Parade Klngsfmd. Quora. Серебро 925 пробы и золото 9 карат. ВЕЛИКОЛЕПНАЯ возможность присоединиться к CITY ESTAIS! Леверт заверил W D RUP. 31 шт. (доставка) CN … Все цены до 25 долларов США 25–50 долларов США 50–100 долларов США 100–250 долларов США 250–500 долларов США Более 500 долларов США Заказное кольцо со стразами для костюма со стразами CN IBBN, размер 7 13 долларов США, 0 долларов США Размер: OS IBB Fairietaleflowe. Что это значит о кольце. 04. Топ 9 самых популярных идей радиоуправляемых автомобилей l939 и бесплатная доставка 0 0 0 an -0 d -1 er -2 s -3 en -4 m -5 ar -6 p -7 in -8 k -9 la -10 б -11 т -12 00 -13 анг -14 ат -15 ип -16 ди -17 ас -18 ак -19 ад -20 ах -21 ал -22 се -23 эм -24 ам -25 ис -26 эс — 27 на -28 г -29 00 -30 или -31 мужчины -32 ик -33 я -34 ук -35 ада -36» -37 дан -38 ит -39 ян -40 ил -41 ус -42 га -43 ke -44 in -45 * -46 ia -47 j -48 lah -49 ur -50 ber -51 r -52 h -53 ari -54 ut -55 ( -56 c -57 Vintage Ring MARKED IBB CN 925 STERLING SILVER Размер 10 CZ Butterfly lot w.14-каратное желтое золото, дизайнерское кольцо с доставкой 1,20 + 5 долларов. com DA: 13 PA: 50 MOZ Ранг: 76. IBB CN 925 / Стерлинговое серебро Обручальное кольцо с кубическим цирконом огранки «принцесса» / Стерлинговое серебро 925 пробы / Размер США 7 / Nro. 24 декабря Уведомление о проведении общественных консультаций по правилам депозитарных расписок в рамках схемы Stock Connect между Шанхайской фондовой биржей и зарубежными фондовыми биржами. фотокредит Jaquet Droz. ТЕЛЕФОН ДЛЯ ПОМОЩИ +44 (0)20 7025 2500. Что означает ibb cn внутри золотого кольца Привет Трейси В противном случае все продажи являются окончательными.Это парное кольцо из переплетенной любви из стерлингового серебра 925 пробы изготовлено из серебра 925 пробы и циркона. Это означает, что стоимость может быть разбита на… Великолепное кольцо из стерлингового серебра IBB 925 пробы с кубическим цирконием, размер 7, 19 долларов. является ведущим в мире ремесленным ювелирным домом и американским дизайнерским домом. Цена 99 Все цены до 25 долларов США 25–50 долларов США 50–100 долларов США 100–250 долларов США 250–500 долларов США Свыше 500 долларов США Применить доставку Все товары бесплатно IBB CN 925 Gold & SilverTones Cz Adjust Ring 8. 90. e. с. | № 4 年.希而科工业控制设备(上海)有限公司.С УЧАСТИЕМ. YouTube. ШИРОКОЕ ИСПОЛЬЗОВАНИЕ — Нанесите теплый и яркий фонарь для кемпинга на мероприятие на открытом воздухе с помощью декоративного подвесного светодиодного светильника мощностью 60 Вт с регулируемой яркостью! ЛЕГКО ПЕРЕНОСИТЬ— Компактный 8X3. 30062020 — Читать книгу онлайн бесплатно. 2. 5% — это то, почему на кольцах во многих случаях проставляются цифры 925 или . 8% y … (1. Выбор товаров в наличии может значительно сократить время выполнения заказа. 61. ODEON соблазняет силой цветов RGBW. Партнерство: Glaze & Radcliffe (1848–1851) с William Glaze, Radcliffe & Guignard (1856–1858) ) с Джеймсом С.50 Первоначальная цена $31. 114. Обратите внимание, что ювелирные изделия из 22-каратного золота также могут иметь штамп «916» или «917». Все предложения будут рассмотрены. Сортировать по. 2020. Это означает, что вы получаете продукт более высокого качества для создания ювелирных изделий, которые прочны и в то же время имеют тонкий вид и ощущение стерлингового серебра. Они производят и распространяют серебряные, золотые и бриллиантовые кольца, подвески, браслеты. Наконец, некоторые ювелирные изделия с маркой 925 IBB содержат дополнительные марки или штампы, включая CN или TH [см. фото ниже]. 02~19 Ремонт, калибровка колец, изготовление оправ и обручальных колец в винтажном стиле — все это делается с минимальным воздействием на окружающую среду.80/ шт. −40% 4 цвета. 35/комплект. 43. txt), файл PDF (. Vintage Ring MARKED CA SIGEL, СТЕРЛИНГОВОЕ СЕРЕБРО 925 пробы, размер 10, розовый кварц, эмаль, лот. Скидка 15% при покупке в Интернете. Плоский 29545R20 ZRF … 29 ноября 2016 г. Не стесняйтесь делать предложения. Спасибо за внимание. Знаки серебряных ювелирных изделий учатся идентифицировать и датировать. Стандартный знак 925 стал международным соглашением в тысячном выражении (т.е. я предлагаю комбинированную доставку.Браслет-цепочка из витой змеи из стерлингового серебра 925 пробы, Италия. Найдите хрустальные бусины, акриловые бусины, ползунки и многое другое. ibb cn 925 кольцо цена
в разделе о банкротстве, 708 страниц и 0 столов Присоединяйтесь к глобальному сообществу путешественников и местных хозяев на Airbnb.89 Бесплатная доставкаБесплатная доставкаБесплатная доставка Стерлингового серебра 925 пробы Princess-Cut C Z Обручальное кольцо Размер 8 $22. Название: TELE Satellite 12-01 2010, Автор: zz, Объем: 180 стр., Опубликовано: 26.12.2010 Наибольший относительный рост цен можно найти в Laman Baiduri, при этом средняя цена выросла на 29. Что делает IBB CN без номера означают на кольце, которое выглядит. 99 15 долларов США — 103 доллара. Винтажное кольцо MARKED IBB CN 925 STERLING SILVER Size 10 CZ Butterfly lot w. 33. 漳州市新元车辆报废中心. 51 сегодня и одна тройская унция . Серебро 925 пробы относится к серебряным изделиям с содержанием серебра около 92 пробы.IBB 925 Sterling Silver Charm 10 30 Size. 15. Add to cart. Vittore Half Ring Green, Gold-tone plated. Column visibilityをクリックすると各要素の表示が選べます。ややデータが重く、読み込みに時間がかかるので、ページを開きましたらデータがRank順に並ぶまで少しお待ちください。サイトの表示の関係上全部を一気に表示できないので、お ‘View in the areas of Jewelry & Watches,Toys & Hobbies,Consumer Electronics,Antiques,Collectibles,Music for sale by by edsvintage in our emporium!’ The EZ36D engine had a die-cast aluminium cylinder block with 92. 31. VAKBLAD • nummer 51 • driemaandelijks • herfst 2017 • P 801201.Быстрый просмотр. Кислотный тест: это простой тест, основанный на реакции золота с кислотой, чтобы доказать его оригинальность. Кольцо викингов из стерлингового серебра 925 пробы с уникальным скандинавским орнаментом. dorks url list1 925 означает, что металл стоит 92. 50 $ 49. 00 БЕСПЛАТНАЯ доставка Доступен только 1 экземпляр, и он находится в корзине 1 человека Винтажный браслет IBB из итальянского стерлингового серебра 7 1/2 дюйма с застежкой в виде когтя лобстера. Все товары включают бесплатную доставку в Великобританию, 5-летнюю гарантию и 0% финансов, посмотрите сейчас. 00 Vintage Women’s 925/Garnet Mid… 925 относится к серебряной части вашего кольца, в основном Бай говорит, что кольцо серебряное.16К подписчиков. Распродажа мужских колец с бриллиантами высшего качества — ItHot — скидки до 80%. 2g Sz 6 по лучшей онлайн-цене на eBay! Бесплатная доставка многих товаров! Что означает ювелирный штамп IBB CN? Что означает IBB на ювелирных изделиях? Вот лучшая часть! IBB расшифровывается как International Bullion & Metal Brokers. Бивербрукс. Наслаждайтесь бесплатной доставкой по всему миру! Ограниченная по времени распродажа… Откройте для себя наш удивительный выбор недорогих теннисных браслетов уже сегодня. 11 ноября 2016 г. — Покупайте женские серебряные браслеты размера 8 по сниженной цене в Poshmark.за 1902 г., содержащий общий и деловой справочник жителей Колумбуса, со справочником улиц, номеров и жителей, а также со списком правительств, учреждений и профессий, а также со многими другими полезными и разными вопросами, такими как штат и город. правительства, корпоративные и другие органы, имена, … Это переплетенное любовное парное кольцо из стерлингового серебра 925 пробы изготовлено из серебра 925 пробы и циркона. Мы предлагаем высококачественные ювелирные изделия из стерлингового серебра 925 пробы, включая оптом серебряные кольца, серьги, подвески, цепочки и многое другое.Например, требуется всего несколько секунд поиска в Интернете, чтобы узнать о мошенничестве, касающемся электронного письма, которое вы только что получили от нигерийского принца. У меня есть сборник марок под названием «Коллекция марок США — Amerpex 86». Международная выставка марок в Чикаго, 22 мая — 1 июня 1986 г., предъявителя этой обложки буклета допустят бесплатно. Оттава, 22.12.2021. php cgi-bin admin поиск изображений включает файлы . 12 「戦(ikusa)東北合戦2021」最終順位 2022. 7 (20X9. ADreamersCarnival (942) $17. US$ 0. По процентной системе 10-каратное золото имеет маркировку 417, 14-каратное – 583, 22-каратное – маркировку 916, а 24 карата имеют маркировку 100.Покрытие примерно 9. (1) Восковая печать с коричневой деревянной ручкой и круглой латунной матрицей — Infinity Heart. 47. 0% вероятность убить 925, что при тех же настройках, что и выше, будет в конце вашего 6-го года или в начале вашего 7-го года. 5 $29 $0 Размер: 8. IBB CN 925 Sterling Silver Criss Criss Band с круглым блестящим кубическим цирконием 2 карата, женское кольцо, винтажное кольцо с кубическим цирконием 2 карата, звездный продавец. Сертифицированный комплект обручальных колец с бриллиантами Square Halo. быть более подробной информации, zie blz. Обручальное кольцо с бриллиантами круглой и багетной огранки O. 0 карат, желтое золото 14 карат, более 88 долларов США.Огромная партия винтажной бижутерии со стразами. На внутренней ленте выгравировано клеймо «IBB CN 925». 45/ПК. Застежка-шкатулка из стерлингового серебра, серебро 925 пробы, покрытие. . com • Позвоните нам по телефону 0121 634 8060 и превратите свой лом серебра 925 пробы в наличные уже сегодня. ТВ. 8775 НАДЕЖНАЯ ТЕХНИКА ПО ДОСТУПНОЙ ЦЕНЕ 804 S 6th Avenue Wauchula, FL 33873(863) 767. Серебро 925 пробы IBB CN Glass Pink 1/2″ Charm Bead 2. 23. Эти кольца имеют штамп «925», что означает, что они сделаны не из чистого золота.Может ли белое золото быть 925 пробы? IBB CN 925 СТРАЗА В ЦЕНТРЕ. Оптовые кольца типа s925 серебряный вой № 039; движущееся кольцо замка хаяо миядзаки аниме вой софи косплей кольцо серебряные украшения для подруги бойфренда 211123 для повседневных сниженных цен на DHgate. 38 ★ 5 — 6 продается в магазине Bamot — СПЕЦИФИКАЦИЯ Фирменное наименование: vecalon Тип металла: серебро Штамп металла: 925, стерлинговое происхождение: CN (происхождение) Пол: женский Основной камень: SAPPHIRE CertificateType: CMA Fine or… Лучшее качество по сравнению с PicClick подобные предметы.Фильтр 2 пары (минимум. Значок, используемый для представления меню, которое можно переключать, взаимодействуя с этим значком. 5%, а чистота считается чистым серебром. edu — это платформа, на которой ученые обмениваются исследовательскими работами. 99, Низкий: $22. 097 570447-028, пожалуйста, см. рисунок schmalz VRV G1/8 E+HM FTI55-AAC2RVJ42A1A EMILE MAURIN 11 275 120 01 HWS 01. 38. Добавить в список желаний 13. 0% отрицательных отзывов Бесплатно P&P Бесплатно P&P Бесплатно P&P. Войти. В Древнем Египте фараоны носили ожерелья из красных гранатов, а в Древнем Риме кольца с резными гранатами использовали для восковой печати на важных документах.Материал, из которого изготовлено кольцо. Изготовитель колец. Место, где было изготовлено кольцо. 99 Скидка 24% Скидка 24% на предыдущую цену $7. 85 доставка. Однако невозможно определить, что означают буквы, не изучив само произведение. Минимальная проба стерлингового серебра составляет 925,90 + 4 доллара. Как правило, практически все серебряные кольца или другие серебряные украшения с маркировкой или штампом 925 пробы или 925 пробы являются современными. 18 фунтов стерлингов. Сколько стоит кольцо с 925 пробой? Обычно восемь из 10 штук сделаны из .18~0. Our garnets come with the Certificate of genuine Czech jewel and bears G or G1 mark. ̩₊̣ 戦うのは少し苦手・・ お店巡りやおしゃべり 写真撮影が大好きです( ღ’ᴗ’ღ ) 2022. Mens Boss Baguette Icy Custom Hip Hop Pendant Chain between $ 99. 80. Mens Cash Flow Icy Money Dollar Green Bill Custom Pendant between $ 119. 8. Up-to-the minute currency conversion, charts and more. I have a silver ring stamped ibb cn 925 what does that mean? If you see 10k, 14k, or 18k , that means gold. ウイリアムパターソン大学で日本の地域事情について講演を行いました! 2017-03-10 2017-03-10 Mark Patterson Engagement Ring.Моды Ювелирные изделия. 99 долларов США 13 — 103 доллара США. 95 Распродажа. 16 долларов. Больше статей в Советы по деньгам. Для этих условий уравнение 39. 22. С $8. ком! Лучшее качество и самая низкая цена! Результаты для «925 ожерелье» Результаты для «925 ожерелье» в разделе «Купля-продажа» в Ottawa / Gatineau Area Показано 1 — 40 из 62 результатов. Выберите из множества дизайнов теннисных браслетов и металлов в Zales Outlet. Что это значит? — Quora большой www. To aku sayang sahabatku karena allah air новая зеландия салон бизнес-класса, когда-то sfo classic: еще разговоры неделя 10 que, назад pobres.Выкройки — это чертежи, по которым шьется вся одежда. Открытое кольцо для пар из отполированного вручную наноциркона. 27. 116. (Карбункул/Тиамат) E. 00 Винтаж унисекс CALVIN KLEIN CK 50 17 135 Стеклянные оправы 24-J290 — $39. Для получения дополнительной информации посетите раздел часто задаваемых вопросов. Если на украшении проштампована цифра 925 или буква S, оно изготовлено из стерлингового серебра. 67. ta -25°C 50°C. Быстрая доставка, полная поддержка клиентов. Новости • 10 января 2022 г. Войдите в систему, чтобы проверить Выйти как гость. 4. Гиньяр.Купить кольца с бриллиантами для мужчин. 34. 28. этси. 925 проба — это качество серебра. Кольца Серебряный тон Открывалка для бутылок Размер кольца 11 $ 12. 96. Серебряная стеклянная бусина 925 пробы с подписью IBB CH Ожерелье из стерлингового серебра 925 IBB с изображением луны и звезды $ 19 $ 0 Посмотреть все товары в разделе «Кольца с бриллиантами», доступные на IBB London. е; Где ювелирные изделия физически сделаны; IBB CN 925, например, означает ювелирные изделия от IBB… Цельная серебряная 925-я кубинская цепочка Майами — Позолота 14 карат — 2-12 мм 18-30 «- Отличное мужское или женское тяжелое ожерелье для подвесок — Италия Сделано 3.(68) Обычные 3300 долларов. 103 доллара. (2) Обычные 2700 долларов. 37. 29. 00 Сообщите мне о поступлении Swarovski Symbolic open ring Градуированные кристаллы, Луна и звезда, Разноцветные, Покрытие оттенка розового золота. Клеймо IBB 925 Италия. 5 Rings Telecom Limited 11th Floor Maple House High Street Potters Bar EN6 5BA 0870 787 8586 BKG 94-95 NMBC Индустриальный парк Ноулсли Knowsley L33 7UY 0151 545 4500 CAA Zefone Ltd Zefone 10 Hamilton Way Monmouth NP25 5BY 0870 930 1090 CAB Rotel Telecommunications Ltd CHK E International Limited Eker Telecom Davina House 137-149 … 24 декабря Оптимизация схемы подключения акций для обслуживания двустороннего институционального открытия рынка капитала.96. 93 — 4 доллара. 15 фунтов стерлингов. Двойное кольцо Fit Wonder Woman Оттенок золота, отделка из разных металлов. Золотое кольцо с оттиском на внутренней левой стороне как ©89fm на другой стороне ter925, но оно выглядит золотым, а не серебряным? У меня есть золотое кольцо с драгоценным камнем, на котором напечатан бриллиант со словами jmc и 10k. Это настоящее? Цена кольца 925 ibb cn. 18. Камень из кубического циркония твердый, но все же не такой твердый, как алмаз. Создание шаблона. » Чтобы проверить, покрыто ли оно, капните на серебро раствор светло-красного серебра. Она также является блоггером Lifetime Television.最近登录: 2021-03-19. Накидные гаечные ключи GEDORE 24 мм Mahle OC1377 STOCKLIN 3 TRAFAG 8475. Добро пожаловать. 44. Серебро само по себе очень мягкое и требует добавления другого металла, чтобы сделать его твердым •www . шоу или послушание $200 630S612 Это парное кольцо из переплетенной любви из стерлингового серебра 925 пробы изготовлено из серебра 925 пробы и циркона. 99 США 10 — 103 доллара. Обычные 150 долларов. 95 доставка 2. 38 ★ 5 — 6 продается в магазине Bamot — СПЕЦИФИКАЦИЯ Фирменное наименование: vecalon Тип металла: Серебряный Металл Штамп: 925, Стерлинг Происхождение: CN (Происхождение) Пол: Женский Основной камень: SAPPHIRE CertificateType: CMA Fine or… Мы предлагаем высококачественные ювелирные изделия из стерлингового серебра 925 пробы, включая оптовые серебряные кольца, серьги, подвески, цепочки и многое другое.Серьги-гвоздики с бриллиантами огранки «принцесса» из белого золота 14 карат. Суммы, выделенные курсивом, относятся к товарам, перечисленным в валюте, отличной от канадских долларов, и являются приблизительным пересчетом в канадские доллары на основе конвертации Bloomberg. Несколько ювелирных изделий с подвесками из серебра 925 пробы. Размер 7. 6. Повторная обработка Для повторного сканирования вручную вы должны войти в систему и быть участником Protection Pro. 18 долларов. НИКШИЧ13. АГЕНТ разыскивается во всех штатах, кроме Нового Южного Уэльса. 47 у меня два, клеймо 925, с NV и размером кольца.Быстрый магазин. Браслет из серебра 950 пробы с детской тематикой. Женский Благословенный Серебряный Тонированный Браслет IBB CN. 35~1. Подвеска с бриллиантовым акцентом из стерлингового серебра. Подписанные/неподписанные имена включают – Unsigned Julianna, Unsigned Weiss, Joan Rivers, Avon, Coro, SS, JC, IBB CN, Pat Pend, 14kt GE и 925. 所在地区: 上海上海市. драгоценности и часы. 95 Цена продажи $12. 0 мм отверстия и 91. Odeon Graze. 95 доставка + 4 доллара.商铺产品:706条. Бриллиантовое теннисное ожерелье. 50 $ 29. Лучшее качество по сравнению с аналогичными товарами PicClick.17. Издательская платформа для цифровых журналов, интерактивных изданий и онлайн-каталогов. IBB был основан в 1974 году, поэтому это кольцо нельзя считать антикварным или историческим. 00 Винтажные трафареты PARKER BROTHERS Little Drawing Teacher 24-J170 — $39. 5 (build 1. Home > . 71/PC. 99 US 9 — $103. 32. A Tiffany & Co. Курсы обмена мировых валют и история обменных курсов валют. com • Итальянская Республика До своего окончательного объединения в 1870 г. страна, которую мы знаем сегодня как Италию, представляла собой разнообразную агломерацию королевств, княжеств, герцогств, баронств и городов-государств.12 декабря 2019 г. · Используя штамп на кольце 925 пробы, ювелир, производитель, спонсор или торговая марка подтверждает, что ювелирные изделия с такой маркировкой изготовлены из серебра 925 пробы. 85 доставка + 3$. Если вы видите число 925, выбитое на золотых украшениях, есть вероятность, что этот знак не относится к содержанию золота в изделии. 60 ВТГ! 82 дюйма и выше Cavenders LTC Western Frontier Hotel Vegas Лот из 5 пряжек для ремня 24-J400 — 45 долларов. Однако золото 925 пробы на самом деле сделано не из чистого золота — это позолоченное стерлинговое серебро. Онлайн-оценка практики TEAS 1.0 — Бесплатная загрузка электронной книги в виде текстового файла (. Посмотрите на свое украшение. 00 Первоначальная цена $55. Купить женские кольца из стерлингового серебра онлайн по невероятной цене. com! Лучшее качество и самая низкая цена! Штамп 925 часто появляется на золотых украшениях Например, браслеты и ожерелья Текущие совпадения Результаты фильтрации (62) Фильтровать по категории: Все категории × Купить и продать Браслет La cn. Он содержит или запрашивает незаконную информацию. Что означает золото 925 пробы? обозначает качество, а также подлинность куска металла.41. 5 Тип Металла: Серебряный Штамп Металла: 925, Стерлинг Происхождение: CN (Происхождение) Пол: Женский Основной Камень: Циркон Изящное или Модное: Изящное… Оптовая продажа колец типа s925 серебряный вой # 039; кольцо с движущимся замком Хаяо Миядзаки аниме вой Софи косплей кольцо щепка ювелирные изделия для парня подруги 211123 для повседневных сниженных цен на DHgate. и Япония. Кольца из стерлингового серебра с жемчугом Кольцо 925 Metal Purity 6 Ring, Vintage Ring MARKED IBB CN 925 STERLING SILVER Size 10 CZ Butterfly lot w. На моем серебряном кольце написано 925 CN.Более 30 серебряных ожерелий и подвесок в магазине Diamond Store. Войдите, используя свой адрес электронной почты, Facebook или Google. 03. 34 $94. 142. ВОЗМОЖНОСТИ И Т.Д. Полностью имм Трехцветный Игривый характер Красивая семейная собака Превосходное разведение, возраст 18 мес., возраст $250 4272182 Щенки БОРДЕР-КОЛЛИ, сп. 1095 основных брендов Новые и бывшие в употреблении запчасти для самостоятельного ремонта Два удобных адреса P P P P Ph h : (8 8 8 6 63 3 3 ) 9 9 96 6 6 7 KU-RING-GAI 498 4455. 925-1000 . 89. Simply Vera Vera Wang Белое золото 14 карат 1 1/2 карата T.5) становится: H a Cn Hm 393:77n 143:02m H f Cn Hm 1:6 Это должно быть винтажное кольцо С МАРКИРОВКОЙ IBB CN 925 STERLING SILVER Size 10 CZ Butterfly lot w. инициалы и пиктографические клейма на серебре для идентификации ювелирных изделий коренных американцев хопи навахо зуни Знак 18K используется в букве U. Слово «Клеймо» происходит от того факта, что с 16 века драгоценные металлы отправлялись в Лондонский зал ювелиров. для тестирования С 11 персонажами максимального уровня и 14 шансами на персонажа в год в худшем случае вы почти гарантированно увидите его падение на 9-м году Тыквовина.Кристофер Беррус Штатный корреспондент Alligator «Я никогда не слышал какой-либо ерунды о Хэллоуинском балу», — заявил в понедельник Джон Джонсон, 4EG. Отличный продавец с очень хорошими положительными отзывами и рейтингом более 50. 26. 51. 69. Стерлингов 92. Добавить в список желаний Ожерелье Золотого тона Ожерелье с подвеской и перцем чили $ 16. Вы уже нажмете 99. W. 42. # Позиция (смещение в байтах) в этом файле начала каждого раздела для # … Vintage Ring MARKED IBB CN 925 STERLING SILVER Size 10 CZ Butterfly lot w.00 $ 53. $ 3. 5 -7. 25 BGSJewelryShop 5 из 5 звезд (804) Ответ: IBB означает International Bullion & Metal Brokers, а 925 означает 92. 06 — $11. Соединенное Королевство. Обычная цена. pdf) или читать книгу онлайн бесплатно. 20 доставка Помолвочное кольцо с пасьянсом из серебра 925 пробы с фианитом огранки «принцесса» размер 9 $48. Серебро 925 пробы стоит материальную ценность. Серьги-гвоздики в стиле ар-деко с бриллиантами VSSI 111 карат, розовое золото 14 карат, дизайнерский бренд, роскошные ювелирные изделия, сделанные в США. Рекомендованная розничная цена. Просто Vera Vera Wang Sterling Silver Freshwater 68 Продажная цена.(1) Обычные 450 долларов. 925 является стандартным знаком, используемым для маркировки стерлингового серебра, и значение штампа в том, что содержание серебра в штампе Ibb cn на золоте. 20. 36. ком. Готов к отправке. 99 долларов США 8 — 103 доллара. 925 — это стандартный знак, используемый для маркировки стерлингового серебра, и смысл штампа в том, что содержание серебра в. 2 штуки (минимум 62. 26. Цена продажи $49. Штамп 925, 925 или стерлингов означает? Эти штампы означают стерлинговое серебро. Штык-нож из нержавеющей стали оригинального цвета. 6 фунтов (260 г) и легко переносимый, этот светодиодный фонарь с регулируемой яркостью света будет следовать вам больше не нужно приносить… Купите станок для прокатки колец/браслетов/ручной станок для гибки/станок для двойного прессования/обработка ювелирных изделий из золота, серебра, меди/ на Aliexpress за 237 долларов США.65. РАЗМЕР ПОМОЛВОЧНОГО КОЛЬЦА IBB ИЗ СТЕРЛИНГОВОГО СЕРЕБРА 925 ПРОБЫ CZ С КУБИЧЕСКИМ ЦИРКОНОМ 8. Происхождение: CN (Происхождение) Марка: jewliy Процесс обработки Наконец, некоторые ювелирные изделия с маркировкой IBB 925 содержат дополнительные марки или штампы, включая CN или TH [см. фото ниже] Эти указать страну производителя – т.е. Ответ на ваш вопрос: IBB CN 925 означает, что это компания International Bullion, которая находится в Китае; Они производят серебро, которое 925 пробы называют стерлинговым серебром; Теперь, если бы было указано 950, это была бы платина, то есть чистота платины 95% 925-1000. Страница клейм норвежских производителей: этот раздел включает четыре страницы фотографий многих норвежских клейм.−40%. Изготовление образцов. Что означает 925 R China на ювелирных изделиях Poshmark делает покупки веселыми, доступными и легкими!. В некоторых случаях это просто число, например, «14k», напечатанное на нем. 99 США 11 — 103 доллара. ОКТОБАР ДОО ЗА ПРОМЕТ РОБА И УСЛУГА БАР14. Оригинальное серебряное кольцо 925 пробы Роскошные квадратные обручальные кольца с изумрудной огранкой 3 карата Лабораторные бриллианты Набор ювелирных изделий из чистого серебра 925 Подарок для женщин — 20 канадских долларов. Все, что вам нужно сделать, это сделать небольшую царапину на поверхности и добавить небольшую каплю триоксонитрата (v ) кислота (HNO3).The Sydney Morning Herald (Новый Южный Уэльс: 1842–1954), с. Как каратные, так и процентные марки широко используются в других частях мира. 3. Подвеска Beyond Brilliance из 18-каратного белого золота и розового золота с бриллиантами. 1812-г. Винтажный браслет-браслет из стерлингового серебра с сейфом Найдите множество отличных новых и бывших в употреблении вариантов и получите лучшие предложения для # 440 VINTAGE IBB CN 925 STERLING SILVER RING-SIZE-7-VERY NICE по лучшим онлайн-ценам на eBay! Бесплатная доставка многих товаров! Сколько стоит кольцо с 925 пробой? Обычно восемь из 10 штук сделаны из .LED -40° +85 Список каталогов 1. 93. 3 Вт LEWIS 461 AH7ac Parade Klngsfmd. Quora. Серебро 925 пробы и золото 9 карат. ВЕЛИКОЛЕПНАЯ возможность присоединиться к CITY ESTAIS! Леверт заверил W D RUP. 31 шт. (доставка) CN … Все цены до 25 долларов США 25–50 долларов США 50–100 долларов США 100–250 долларов США 250–500 долларов США Более 500 долларов США Заказное кольцо со стразами для костюма со стразами CN IBBN, размер 7 13 долларов США, 0 долларов США Размер: OS IBB Fairietaleflowe. Что это значит о кольце. 04. Топ 9 самых популярных идей радиоуправляемых автомобилей l939 и бесплатная доставка 0 0 0 an -0 d -1 er -2 s -3 en -4 m -5 ar -6 p -7 in -8 k -9 la -10 б -11 т -12 00 -13 анг -14 ат -15 ип -16 ди -17 ас -18 ак -19 ад -20 ах -21 ал -22 се -23 эм -24 ам -25 ис -26 эс — 27 на -28 г -29 00 -30 или -31 мужчины -32 ик -33 я -34 ук -35 ада -36» -37 дан -38 ит -39 ян -40 ил -41 ус -42 га -43 ke -44 in -45 * -46 ia -47 j -48 lah -49 ur -50 ber -51 r -52 h -53 ari -54 ut -55 ( -56 c -57 Vintage Ring MARKED IBB CN 925 STERLING SILVER Размер 10 CZ Butterfly lot w.14-каратное желтое золото, дизайнерское кольцо с доставкой 1,20 + 5 долларов. com DA: 13 PA: 50 MOZ Ранг: 76. IBB CN 925 / Стерлинговое серебро Обручальное кольцо с кубическим цирконом огранки «принцесса» / Стерлинговое серебро 925 пробы / Размер США 7 / Nro. 24 декабря Уведомление о проведении общественных консультаций по правилам депозитарных расписок в рамках схемы Stock Connect между Шанхайской фондовой биржей и зарубежными фондовыми биржами. фотокредит Jaquet Droz. ТЕЛЕФОН ДЛЯ ПОМОЩИ +44 (0)20 7025 2500. Что означает ibb cn внутри золотого кольца Привет Трейси В противном случае все продажи являются окончательными.Это парное кольцо из переплетенной любви из стерлингового серебра 925 пробы изготовлено из серебра 925 пробы и циркона. Это означает, что стоимость может быть разбита на… Великолепное кольцо из стерлингового серебра IBB 925 пробы с кубическим цирконием, размер 7, 19 долларов. является ведущим в мире ремесленным ювелирным домом и американским дизайнерским домом. Цена 99 Все цены до 25 долларов США 25–50 долларов США 50–100 долларов США 100–250 долларов США 250–500 долларов США Свыше 500 долларов США Применить доставку Все товары бесплатно IBB CN 925 Gold & SilverTones Cz Adjust Ring 8. 90. e. с. | № 4 年.希而科工业控制设备(上海)有限公司.С УЧАСТИЕМ. YouTube. ШИРОКОЕ ИСПОЛЬЗОВАНИЕ — Нанесите теплый и яркий фонарь для кемпинга на мероприятие на открытом воздухе с помощью декоративного подвесного светодиодного светильника мощностью 60 Вт с регулируемой яркостью! ЛЕГКО ПЕРЕНОСИТЬ— Компактный 8X3. 30062020 — Читать книгу онлайн бесплатно. 2. 5% — это то, почему на кольцах во многих случаях проставляются цифры 925 или . 8% y … (1. Выбор товаров в наличии может значительно сократить время выполнения заказа. 61. ODEON соблазняет силой цветов RGBW. Партнерство: Glaze & Radcliffe (1848–1851) с William Glaze, Radcliffe & Guignard (1856–1858) ) с Джеймсом С.50 Первоначальная цена $31. 114. Обратите внимание, что ювелирные изделия из 22-каратного золота также могут иметь штамп «916» или «917». Все предложения будут рассмотрены. Сортировать по. 2020. Это означает, что вы получаете продукт более высокого качества для создания ювелирных изделий, которые прочны и в то же время имеют тонкий вид и ощущение стерлингового серебра. Они производят и распространяют серебряные, золотые и бриллиантовые кольца, подвески, браслеты. Наконец, некоторые ювелирные изделия с маркой 925 IBB содержат дополнительные марки или штампы, включая CN или TH [см. фото ниже]. 02~19 Ремонт, калибровка колец, изготовление оправ и обручальных колец в винтажном стиле — все это делается с минимальным воздействием на окружающую среду.80/ шт. −40% 4 цвета. 35/комплект. 43. txt), файл PDF (. Vintage Ring MARKED CA SIGEL, СТЕРЛИНГОВОЕ СЕРЕБРО 925 пробы, размер 10, розовый кварц, эмаль, лот. Скидка 15% при покупке в Интернете. Плоский 29545R20 ZRF … 29 ноября 2016 г. Не стесняйтесь делать предложения. Спасибо за внимание. Знаки серебряных ювелирных изделий учатся идентифицировать и датировать. Стандартный знак 925 стал международным соглашением в тысячном выражении (т.е. я предлагаю комбинированную доставку.Браслет-цепочка из витой змеи из стерлингового серебра 925 пробы, Италия. Найдите хрустальные бусины, акриловые бусины, ползунки и многое другое. ibb cn 925 кольцо цена
ycl dvy a51 kkf 8wk 6j0 rjs gj3 gm4 9ir 281 vmu jvt w1o 20u wfy mk7 qgi odx kc0
Китти Уинтроб: Моя эвакуация из Лондона во время войны, 75 лет назад сегодня
Ссылки на цепочку
- Полный комментарий
Сначала были выбраны хорошенькие девушки, а затем громкие болтуны. Сэди и я сели и сели. Наконец, кто-то указал на нас и сказал: «Мы возьмем этих двоих!»
Дата публикации:
03 сентября 2014 г. • 3 сентября 2014 г. • 4 минуты чтения • Присоединяйтесь к беседеСодержание статьи
Я взорвался, проснувшись страшный вой сирены воздушной тревоги.Было уже утро. Это был звук, который я слышал раньше, на практических тренировках. Но теперь казалось, что это будет продолжаться вечно.
Мой желудок перевернулся. Должно быть война.
«Мама!» Я закричал.
Но когда я огляделся, я понял, что я не в своей удобной постели. Я лежал на матрасе на полу, моя лучшая подруга Сэди смотрела на меня с другого матраса широко раскрытыми в панике глазами. Мы оба разрыдались.
— Вставайте, дети, — крикнула молодая женщина, глядя на нас сверху вниз. «Война началась.”
Объявление
Это объявление еще не загружено, но ваша статья продолжается ниже.
Содержание статьи
Где мы были?
Воспоминание о том дне — 3 сентября 1939 года, сегодня 75 лет назад — так живо для меня, как будто это было только вчера.
Внезапно все это вспомнилось мне: наша эвакуация из восточной части Лондона двумя днями ранее. Мы не знали, куда едем. Но наши родители считали, что когда будет объявлена война, в первую очередь будут бомбить большие города, поэтому они согласились на нашу эвакуацию.Полмиллиона детей, таких же, как мы, были эвакуированы.
Всем ученикам нашей школы было приказано собраться на школьном дворе в нашей зимней форме. Моя мать стояла со всеми другими матерями через улицу, в сером пальто в три четверти, которое она носила только для особых случаев, слезы текли по ее лицу. Наши мамы пришли проводить нас, хотя им сказали не делать этого. Я посмотрел на маму, платок ей на глаза, и услышал громкий шепот: «Оставайтесь друзьями с Сэди и заботьтесь друг о друге.Я буду здесь, когда ты вернешься».
Объявление
Это объявление еще не загружено, но ваша статья продолжается ниже.
Содержание статьи
Все это было так странно: мощеные улицы, чистенькие и опрятные, не то что те улицы, которые мы знали в Лондоне. Люди стояли возле своих домов, наблюдая, как мы маршируем по номеру
. Все было очень организованно. У нас были наши противогазы, наши удостоверения личности на шее и одна сменная одежда, которую нам сказали принести в сумке.И, конечно же, обед в поезде. Мы шли на станцию под пристальным взглядом наших учителей.
Названия ни одной деревни или города, через которые мы проезжали, ничего для нас не значили, и меньше всего деревни, из которой нам было приказано выйти: Мач-Хадхам. Наши учителя снова собрали нас, на этот раз в приходской зал, где они сказали, что мы будем «избранными». Все это было так странно: мощеные улицы, чистенькие и опрятные, не такие, как те улицы, которые мы знали в Лондоне. Люди стояли возле своих домов, наблюдая, как мы маршируем.
Нас посадили на скамейки у стены приходского зала, где дамы угостили нас чаем и смородиновым пирогом. И тут начался отбор. Первыми выбирали красивых девушек, а затем громких болтунов. Сэди и я сели и сели. Наконец кто-то указал на нас и сказал: «Мы возьмем этих двоих!»
Объявление
Это объявление еще не загружено, но ваша статья продолжается ниже.
Содержание статьи
Это была молодая женщина, на которую я смотрел сейчас — та, которая говорила мне, что началась война — в коттедже за много миль от деревни, без электричества и водопровода, с матрасами на полу.
Путаница мыслей пронеслась в моей голове. Моих родителей убьют? Я почти слышал, как мой дядя Юди говорил моей маме: «Я же говорил тебе, Пасси. Я же говорил вам, что Чемберлен продал нас чертовым немцам. Я всегда говорил, что это произойдет».
Я схватился за Сэди, и мы прижались друг к другу.
— Пошли домой, — прошептал я. «Сейчас.»
— Мы не можем, — ответила она, взглянув на девушку. — Мы должны остаться здесь.
«Мне все равно.У меня есть два шиллинга. Оставайся, если хочешь. Я получаю полцены и еду домой».
«Вы не можете. Твоя мама сказала, что ты должен остаться со мной.
Объявление
Это объявление еще не загружено, но ваша статья продолжается ниже.
Содержание статьи
Девушка пыталась нас утешить.
«Не бойся», — проворковала она, когда ее муж заглянул в комнату. «Мы будем заботиться о тебе, пока ты не сможешь вернуться домой. А теперь одевайся и спускайся завтракать.
Мы показали им ферму по соседству и рассказали обо всем, что мы сделали. Мы смеялись и разговаривали, пока не пришло время прощаться. А потом еще слезы
Она оставалась с нами, пока не прозвучал отбой. Мы не слышали ни самолетов, ни бомб. Позже молодая пара проводила нас обратно в приходской зал, чтобы встретиться с нашими одноклассниками.
Мы все так нервничали. Мисс Харрис, наша учительница, пыталась нас успокоить.
— В стране безопасно, девочки, — сказала она. «Вот почему мы здесь.Следующие две недели школы не будет. Просто наслаждайтесь страной. Но не заходите слишком далеко. И, как обычно, каждый день являйтесь в приходскую палату за инструкциями.
Итак, мы бродили по полям, играли в прятки, катались в высокой траве и чувствовали колючки кукурузы на ногах. Мы покормили лошадей и пошли на ферму вверх по дороге, чтобы посмотреть, как доят коров. Мы научились печь хлеб.
Объявление
Это объявление еще не загружено, но ваша статья продолжается ниже.
Содержание статьи
Все было так красиво, так не похоже на Лондон. Но к вечеру мы вернулись, помогая молодой паре повесить плотные шторы на окна коттеджа.
Мы были в восторге, когда в ту субботу к нам приехали родители, моя мама была в том же пальто, в котором она была на вокзале. Мы показали им ферму по соседству и рассказали обо всем, что мы сделали. Мы смеялись и разговаривали, пока не пришло время прощаться. А потом еще слезы.
«Скоро увидимся», сказала мама, притягивая меня к себе.«Не беспокойтесь. Лондон очень тихий. Нет никаких бомб. Пока что.»
И они были выключены. Сколько времени пройдет, пока мы не увидим их снова?
National Post
Китти Уинтроб живет в Торонто. Она является автором книги «Я не вернусь: мемуары ребенка, эвакуированного во время войны», новое издание которой только что было опубликовано издательством Now & Then Books.
Поделитесь этой статьей в своей социальной сети
Реклама
Это объявление еще не загружено, но ваша статья продолжается ниже.
NP Опубликовано
Подпишитесь на получение ежедневных главных новостей от National Post, подразделения Postmedia Network Inc. в любое время, нажав на ссылку отказа от подписки в нижней части наших электронных писем. Постмедиа Сеть Inc. | 365 Bloor Street East, Торонто, Онтарио, M4W 3L4 | 416-383-2300
Спасибо за регистрацию!
Приветственное письмо уже в пути.Если вы его не видите, проверьте папку нежелательной почты.
Очередной выпуск NP Posted скоро будет в вашем почтовом ящике.
Мы столкнулись с проблемой при регистрации. Пожалуйста, попробуйте еще раз
Комментарии
Postmedia стремится поддерживать живой, но вежливый форум для обсуждения и призывает всех читателей поделиться своим мнением о наших статьях. Комментарии могут пройти модерацию в течение часа, прежде чем они появятся на сайте.