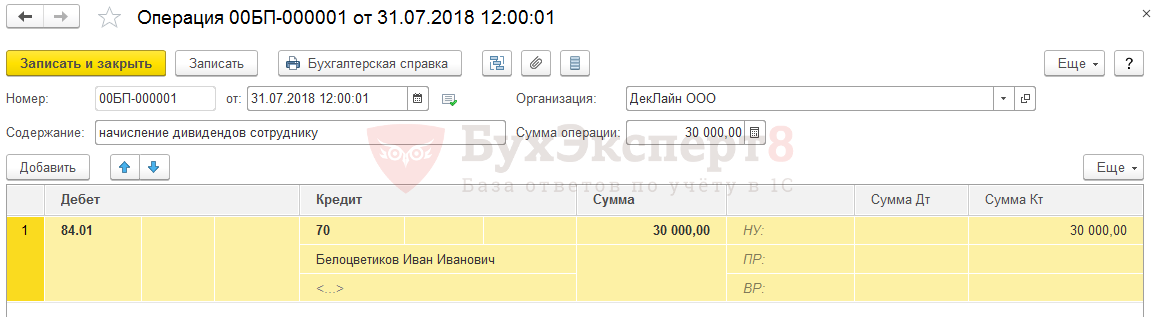
Начисление дивидендов в 2019 году: проводки в бухучете
Как происходит начисление
Дивиденты представляют собой одну из составляющих прибыли, полученной организацией, которая перераспределяется между учредителями. Размер напрямую зависит от доли каждого участника в уставном капитале или пакета акций, которым владеет тот или иной учредитель.
Начисление дивидендов (проводки) производится непосредственно в день распределения полученной чистой прибыли.
Выплачиваются дивиденды (проводки формируются по счету 84) за счет средств нераспределенной прибыли.
Выплаты производятся на основании регламента, принятого и утвержденного в организации. Этот порядок согласуется всеми членами акционерного собрания предприятия, подтверждением чего является подписанный ими акт или решение. Частота определена законодательно (п. 1 ст. 28 14-ФЗ от 08.02.1998) и бывает:
- ежеквартальной;
- полугодовой;
- ежегодной.
Выплаты учредителям
Начислены дивиденды, проводка будет зависеть от того, к какой категории относится получатель.
Если получателем выступает физическое лицо, например, работник организации, то бухучет дивидендов (проводки) формируется следующим образом:
Дт 84 Кт 70.
Для сч. 70 «Расчеты с персоналом по оплате труда» создается субсчет, который и будет отражать все взаиморасчеты с акционерами-сотрудниками. Удержание НДФЛ проводится следующей бухгалтерской записью:
Дт 70 Кт 68.
В том случае, когда участником финансовой доли в организации является юридическое либо физическое лицо, которое не является штатным сотрудником, то отражение в учете будет проводиться следующими операциями:
- Дт 84 Кт 75.2;
- Дт 75.2 Кт 68 — удержание подоходного налога.
Начисления по акциям
Доходность, начисляемая акционерным обществом, может быть распределена между участниками как в натуральной, так и в денежной формах.
Начислены дивиденды, бухгалтерские проводки по акциям:
- Дт 75.2 Кт 50 — выдача в виде наличных средств в кассе организации;
- Дт 75 Кт 90 — доходность по акциям выдана участнику в натуральной форме, отражение стоимости готовой продукции;
- Дт 90 Кт 43 — списание себестоимости по ДВ, выданным в товарной форме.
Как вести бухучет
Бухгалтерские записи будут различны для каждой категории участников-получателей. Все операции должны сопровождаться исчислением и удержанием налоговых платежей, которые проводятся непосредственно в день перечисления.
Для получателей, которые являются сотрудниками организации, выплаты будут проведены следующим образом:
Дт 70 Кт 50, 51.
Прочие участники получают по следующей бухгалтерской записи:
Дт 75.2 Кт 50, 51.
Если часть суммы осталась на счетах организации по причине невостребованности участниками выплаты, то такие финансовые ресурсы подлежат списанию через операцию:
Дт 70, 75 Кт 84.


Бухгалтерский учет у участника — получателя доходов
Если учредителем, получающим доходность в такой форме, выступает юридическое лицо, то ему также надлежит проводить такие поступления. Данный вид платежей приходуется на счете 91 «Прочие доходы и расходы». Согласно п. 7, 16 ПБУ 9/99, доходность, полученная от подобных выплат, принимается к бухучету непосредственно в дату принятия решения о перечислении. В форме № 1 бухгалтерской отчетности (ББ) такие поступления можно посмотреть в строке 2310.
При получении дивидендов учредитель формирует следующие записи:
- Дт 76.3 Кт 91.1 — зачислены в прочие доходы предприятия-участника;
- Дт 50, 51 Кт 76.3 — поступили получателю.
Об авторе статьи
Задорожнева Александра
Бухгалтер
В 2009 году закончила бакалавриат экономического факультета ЮФУ по специальности экономическая теория.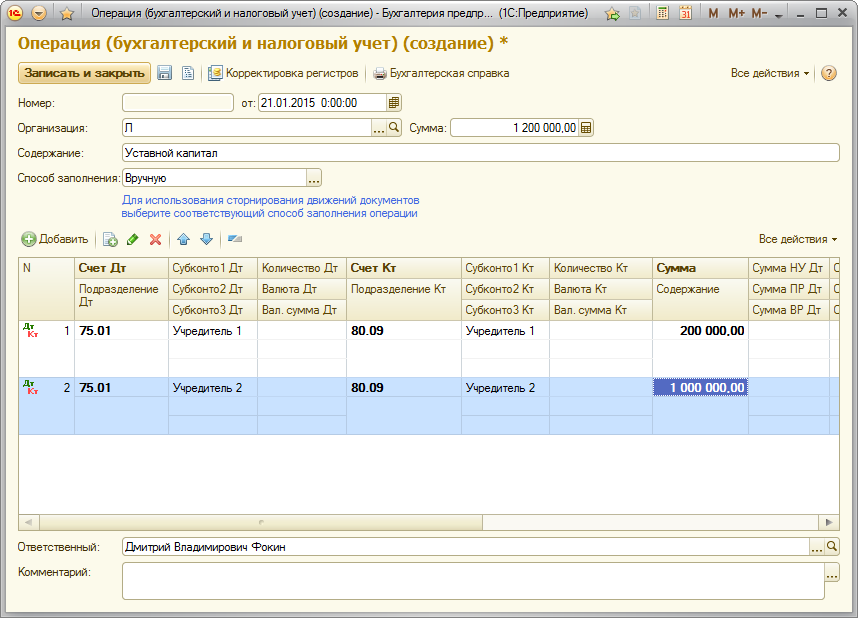 В 2011 — магистратуру по направлению «Экономическая теория», защитила магистерскую диссертацию.
В 2011 — магистратуру по направлению «Экономическая теория», защитила магистерскую диссертацию.
Организация бухгалтерского учета на предприятии. Тема 10. Учет финансовых результатов и нераспределенной прибыли. Тест для самопроверки
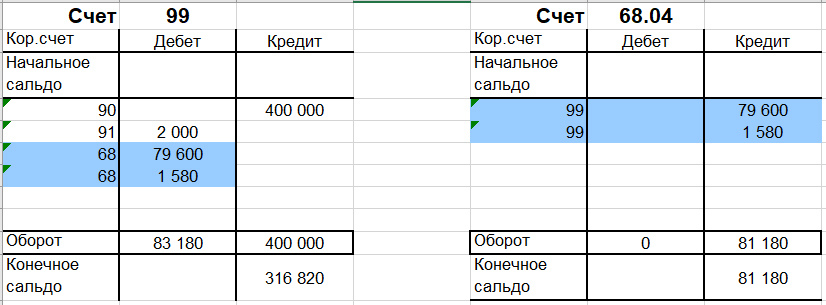
Дт 68 Кт 99
Дт 99 Кт 68
Дт 91 Кт 68
2. Финансовый результат от прочих доходов и расходов определяют на счете …
90
91
99
3. Счет 99 «Прибыли и убытки» …
активный
активно-пассивный
пассивный
4. Чистая прибыль списывается в конце года …
Дт 99 Кт 82
Дт 99 Кт 84
Дт 90 Кт 99
5. Списание потерь материальных ценностей в результате стихийного бедствия
Дт 91 Кт 10
Дт 99 Кт 10
Дт 84 Кт 10
6. Расходы будущих периодов — это …
Расходы будущих периодов — это …
средства, полученные в отчетном периоде, но относящиеся к будущим отчетным периодам
затраты, произведенные организацией в отчетном периоде
затраты, произведенные организацией в предшествующем и (или) отчетном периодах, но подлежащие включению в себестоимость продукции (работ, услуг) в последующие периоды
7. Проводка, которой оформляют реформацию баланса при получении прибыли по итогам года
Дт 84 Кт 99
Дт 83 Кт 99
Дт 99 Кт 84
8. Не может иметь остаток на конец отчетного года резерв на …
выплату ежегодного вознаграждения за выслугу лет
выплату вознаграждений по итогам работы за год
ремонт основных средств (ремонтный фонд)
гарантийный ремонт и гарантийное обслуживание
9. Произведены отчисления от прибыли в резервный капитал
Дт 86 Кт 99
Дт 91 Кт 86
Дт 99 Кт 86
Дт 84 Кт 82
Дт 91 Кт 83
10. Прибыль направлена на выплату доходов учредителям и участникам
Дт 99 Кт 75(70)
Дт 91 Кт 75(70)
Дт 84 Кт 75 (70)
11. Конечный финансовый результат за отчетный период определяют на счете 99 как …
Конечный финансовый результат за отчетный период определяют на счете 99 как …
разницу между кредитовым и дебетовым оборотами
кредитовый оборот
12. Финансовый результат от продажи материалов отражается на счете …
84 «Нераспределенная прибыль (непокрытый убыток)
90 «Продажи»
91 «Прочие доходы и расходы»
99 «Прибыли и убытки»
13. В состав доходов будущих периодов включаются ….
субсидии, полученные из государственного бюджета
арендная плата, полученная авансом
возмещение убытков, полученных в результате стихийного бедствия
14. Финансовый результат (прибыль) от реализации продукции отражают проводкой …
Дт 99 Кт 90
Дт 90 Кт 99
Дт 91 Кт 99
15. Финансовый результат деятельности организации отражается на счете …
84 «Нераспределенная прибыль (непокрытый убыток) »
91 «Прочие доходы и расходы»
99 «Прибыли и убытки»
16.
Дт 91 Кт 51
Дт 51 Кт 91
Дт 99 Кт 51
17. Создание резервов предстоящих расходов …
не обязательно
обязательно
проводится по решению налоговых органов
Учет нераспределенной прибыли.
Для обобщения информации о наличии и движении сумм нераспределенной прибыли или непокрытого убытка организации используют А/П счет 84 «нераспределенная прибыль (непокрытый убыток)».
Сумму чистой прибыли отчетного года списывают заключительными оборотами декабря в кредит счета 84 «нераспределенная прибыль (непокрытый убыток)» со счета 99 «Прибыли и убытки» (счет 99 дебетуется).
Распределение
прибыли осуществляется на основание
решения общего собрания акционеров в
акционерном обществе , собрание участников
в ООО или другого компетентного органа.
На суммы начисленных доходов учредителя – Дт 84 Кт 70, 75. Аналогично запись осуществляется при начислении промежуточных доходов.
Наряду с созданием спец фонда акционирования работников АО могут создавать фонды выплаты дивидендов по привилегированным акциям определенных типов. Учет данного фонда целесообразно осуществлять на субсчете «Фонд на выплату дивидендов по привилегированным акциям» счета 84. Начисления дивидендов по привилегированным акциям за счет специального фонда отражают – Дт 84 субсчет «Фонд на выплату дивидендов по привилегированным акциям» Кт 75.
Начисления дивидендов по обыкновенным акциям – Дт 84 Кт 75, 70.
Отчисления в резервный капитал – Дт 84 Кт 82.
Направления чистой прибыли на покрытие убытка предыдущего года – Дт 84 Кт 84.
Сумма
чистого убытка отчетного года списывается
с закл. оборотами декабря в Дт 84 с Кт 99.
Для обобщения информации о наличии и
движении сумм нераспределенной прибыли
или непокрытого убытка организации
используют активно-пассивный счет 84
«Нераспределенная прибыль (непокрытый
убыток)». Сумму чистой
прибыли отчетного года списывают
заключительными оборотами декабря в
кредит счета 84 «Нераспределенная
прибыль (непокрытый убыток)» со счета
99 «Прибыли и убытки» (счет 99
дебетуется). Распределение
прибыли осуществляется на основании
решения общего собрания акционеров в
акционерном обществе, собрания участников
в обществе с ограниченной ответственностью
или другого компетентного органа. Чистая
прибыль может быть направлена на выплату
дивидендов, создание и пополнение
резервного капитала (фонда), покрытие
убытков прошлых лет. На суммы
начисленных доходов учредителям дебетуют
счет 84 «Нераспределенная прибыль
(непокрытый убыток)» и кредитуют счета
70 «Расчеты с персоналом по оплате
труда» (работникам организации) и 75
«Расчеты с учредителями» (сторонним
участникам).
При размещении дополнительных обыкновенных акций по цене ниже рыночной стоимости. В первом случае каждому акционеру — владельцу обыкновенных акций распределяется целое число обыкновенных акций пропорционально числу принадлежащих ему обыкновенных акций. Для обеспечения сравнимости средневзвешенного количества обыкновенных акций, находящихся в обращении, на начало и конец отчетного периода, обыкновенные акции считаются размещенными на начало отчетного периода. При этом количество обыкновенных акций, находящихся в обращении до даты указанного размещения, при расчете их средневзвешенного количества увеличивается (или уменьшается) в той же пропорции, в какой они были увеличены (или уменьшены) в результате указанного размещения.
12.7. Учет нераспределенной прибыли (непокрытого убытка)
Читайте также
УЧЕТ ИСПОЛЬЗОВАНИЯ ПРИБЫЛИ
УЧЕТ ИСПОЛЬЗОВАНИЯ ПРИБЫЛИ
В соответствии с законодательством предприятия платят из валовой прибыли в бюджет налог на прибыль и отдельные виды доходов, оставшаяся часть (чистая прибыль) используется предприятием. Распределение налогооблагаемой прибыли отражают на
Распределение налогооблагаемой прибыли отражают на
12.7. Учет нераспределенной прибыли (непокрытого убытка)
12.7. Учет нераспределенной прибыли (непокрытого убытка) Для обобщения информации о наличии и движении сумм нераспределенной прибыли или непокрытого убытка на сельскохозяйственных предприятиях используют активно–пассивный счет 84 «Нераспределенная прибыль (непокрытый
89. Анализ нераспределенной прибыли и оценочных резервов
89. Анализ нераспределенной прибыли и оценочных резервов Нераспределенная прибыль – это часть капитала держателей остаточных прав (собственников), аккумулирующая невыплаченную в виде дивидендов прибыль, которая является внутренним источником финансовых средств
Глава 3.
 Формирование прибыли (убытка) организации
Формирование прибыли (убытка) организации
Глава 3. Формирование прибыли (убытка) организации В бухгалтерской и управленческой повседневной практике принято различать финансовые результаты вообще и чистую прибыль в частности, хотя в действительности чистая прибыль является составной частью финансовых
3.1. Формирование чистой прибыли (убытка)
3.1. Формирование чистой прибыли (убытка) Главный финансовый результат, как он понимается в экономической науке, есть прибыль или ее «зеркальное отражение» – убыток, полученные в результате оборота капитала за отчетный период. Прибыль с незапамятных времен (еще с XV в.,
12.1. Учет формирования прибыли
12.1. Учет формирования прибыли
12. 1.1. Какие нормативные документы определяют порядок учета формирования и распределения прибыли?? Налоговый кодекс Российской Федерации.? Положение по бухгалтерскому учету «Доходы организации» (ПБУ 9/99), утвержденное приказом Минфина
1.1. Какие нормативные документы определяют порядок учета формирования и распределения прибыли?? Налоговый кодекс Российской Федерации.? Положение по бухгалтерскому учету «Доходы организации» (ПБУ 9/99), утвержденное приказом Минфина
13.4. Учет нераспределенной прибыли
13.4. Учет нераспределенной прибыли 13.4.1. Каким образом учитывают нераспределенную прибыль (непокрытый убыток)?Обобщение информации о наличии и движении сумм нераспределенной прибыли или непокрытого убытка предприятия осуществляется на счете 84 «Нераспределенная прибыль
109. Учет прибыли (убытков) от обычных видов деятельности предприятия
109. Учет прибыли (убытков) от обычных видов деятельности предприятия
Доходы от обычных видов деятельности отражаются по счету 90 «Продажи», субсчет 1 «Выручка». К ним относится выручка от продажи продукции и товаров, поступления, связанные с выполнением работ, оказанием
К ним относится выручка от продажи продукции и товаров, поступления, связанные с выполнением работ, оказанием
113. Учет нераспределенной прибыли (непокрытого убытка) предприятия
113. Учет нераспределенной прибыли (непокрытого убытка) предприятия Учет нераспределенной прибыли (непокрытого убытка) ведется на пассивном счете 84 «Нераспределенная прибыль (непокрытый убыток)». Счет 84 предназначен для обобщения информации о наличии и движении сумм
1. Конечный источник прибыли и убытка на рынке
1. Конечный источник прибыли и убытка на рынке Постоянные изменения начальных данных, препятствующие превращению экономической системы в равномерно функционирующую экономику, вновь и вновь порождающие предпринимательские прибыли и убытки, для одних членов
2.
 4. Увеличение уставного капитала за счет нераспределенной прибыли
4. Увеличение уставного капитала за счет нераспределенной прибыли
2.4. Увеличение уставного капитала за счет нераспределенной прибыли Если фирма работает успешно и после уплаты всех налогов у нее остается нераспределенная прибыль, то ее учредители имеют право увеличить уставный капитал общества на сумму этой прибыли.В соответствии с
2.4. Увеличение уставного капитала за счет нераспределенной прибыли
2.4. Увеличение уставного капитала за счет нераспределенной прибыли 2.4.1. Законодательные основы и отражение в учете Если компания работает успешно и после уплаты всех налогов у нее остается нераспределенная прибыль, то ее учредители имеют право увеличить уставный
Вопрос 46 Анализ формирования нераспределенной прибыли
Вопрос 46
Анализ формирования нераспределенной прибыли
Анализ нераспределенной прибыли целесообразно начать с изучения ее состава и динамики изменения отдельных статей. В состав нераспределенной прибыли должны быть включены следующие статьи формы № 2 «Отчет о
В состав нераспределенной прибыли должны быть включены следующие статьи формы № 2 «Отчет о
Промежуточные дивиденды при убытке
Как учитываются промежуточные дивиденды при убытке? Может сложиться ситуация, когда дивиденды выплачены, а по итогам календарного года сформирован убыток. Как быть с выплаченными собственникам суммами? Правильный ответ – в статье.
Бухучет
Сразу скажем, что такие суммы дивидендами считать нельзя. Нет прибыли, нет и дивидендов. Поэтому такие выплаты квалифицируются как безвозмездные. Напомним, что дивиденды выплачивают только из чистой прибыли. В свою очередь, ее размер окончательно определяется только по итогам года. Значит, суммы выплаченные учредителям при отсутствии прибыли признаются безвозмездными выплатами.
Такие «дивиденды» нужно отразить в составе прочих доходов организации:
Дт 91-2 Кт 76 (73) – отражено начисление безвозмездной выплаты учредителю.
Одновременно с учетом расходов нужно сторнировать начисление дивидендов в бухучете:
Дт 84 Кт 75-2 (70) – сторнированы дивиденды учредителя.
Когда списать промежуточные дивиденды при убытке? В момент выявления убытка (п. 11 ПБУ 10/99).
Налоговый учет
Теперь расскажем про налогообложение прибыли в этой ситуации. Безвозмездную выплату в расходах по налогу на прибыль учесть нельзя (п. 49 ст. 270 НК РФ).
Отметим, что компании, выплачивающие дивиденды участникам, должны исполнять обязанности налогового агента. Иными слова, нужно рассчитать, удержать и перечислить налог на прибыль в бюджет с суммы дивидендов, полученных участником. Если окажется, что выплаченные суммы дивидендами не являются, то переплаченную в бюджет сумму можно вернуть, подав в налоговую инспекцию уточненную декларацию.
В свою очередь, участник общества должен будет заплатить налог на прибыль с полученных средств. Рассчитаться с бюджетом придется по общей ставке 20%, вместо 13% (15%), предусмотренных для доходов в виде дивидендов.
НДФЛ
В случае переквалификации дивидендов в безвозмездную выплату организация-плательщик будет вынуждена дополнительно удержать НДФЛ у участников-нерезидентов. Дело в том, что для таких выплат предусмотрена более высокая ставка налога, чем для дивидендов: 30% вместо 15%. Если сделать это не получается, то нужно уведомить налоговую инспекцию по месту учета организации, а также гражданина получившего доход (п. 5 ст. 226 НК РФ). Такие последствия могут быть при выплате промежуточных дивидендов при убытке.
Если речь идет о переквалификации доходов налоговых резидентов России, то удерживать налог не потребуется. Объясняется это тем, что и в первом и во втором случае применяется одинаковая ставка – 13%. (ст. 224 НК РФ, письмо ФНС от 19.03.2009 № ШС-22-3/210).
Увеличение уставного капитала АО за счет дивидендов физических лиц отражается записью Дт 70 «Расчеты с персоналом по оплате труда»
Сумма складочного капитала при создании хозяйственного товарищества отражается записью
(*ответ*) Дт 75 «Расчеты с учредителями» — Кт 80 «Уставный капитал»
Дт 01 «Основные средства», 04 «Нематериальные активы» — Кт 75 «Расчеты с учредителями»
Дт 80 «Уставный капитал» — Кт 75 «Расчеты с учредителями»
Дт 55 «Специальные счета в банках» — Кт 75 «Расчеты с учредителями»
Суммы фактически внесенных денежных средств в счет вклада в уставный капитал АО отражаются записью
(*ответ*) Дт 51 «Расчетные счета» — Кт 75 «Расчеты с учредителями»
Дт 80 «Уставный капитал» — Кт 75 «Расчеты с учредителями»
Дт 75 «Расчеты с учредителями» — Кт 80 «Уставный капитал
Дт 75 «Расчеты с учредителями» — Кт 51 «Расчетные счета»
Увеличение складочного капитала за счет средств от переоценки основных фондов после перерегистрации товарищества отражается записью
(*ответ*) Дт 83 «Добавочный капитал» — Кт 80 «Уставный капитал»
Дт 01 «Основные средства» — Кт 80 «Уставный капитал»
Дт 84 «Нераспределенная прибыль (непокрытый убыток)» — Кт 80 «Уставный капитал»
Дт 82 «Резервный капитал» — Кт 80 «Уставный капитал»
Увеличение складочного капитала товарищества за счет свободного остатка нераспределенной прибыли после перерегистрации отражается записью
(*ответ*) Дт 84 «Нераспределенная прибыль (непокрытый убыток)» — Кт 80 «Уставный капитал»
Дт 01 «Основные средства» — Кт 80 «Уставный капитал»
Дт 83 «Добавочный капитал» — Кт 80 «Уставный капитал»
Дт 82 «Резервный капитал» — Кт 80 «Уставный капитал»
Увеличение складочного капитала товарищества за счет средств резервного фонда после перерегистрации отражается записью
(*ответ*) Дт 82 «Резервный капитал» — Кт 80 «Уставный капитал»
Дт 01 «Основные средства» — Кт 80 «Уставный капитал»
Дт 84 «Нераспределенная прибыль (непокрытый убыток)» — Кт 80 «Уставный капитал»
Дт 83 «Добавочный капитал» — Кт 80 «Уставный капитал»
Увеличение стоимости основных средств в результате их переоценки отражается по кредиту счета
(*ответ*) 83 «Добавочный капитал»
80 «Уставный капитал»
82 «Резервный капитал»
84 «Нераспределенная прибыль (непокрытый убыток)»
Увеличение уставного капитала АО за счет дивидендов физических лиц отражается записью
(*ответ*) Дт 70 «Расчеты с персоналом по оплате труда» — Кт 80 «Уставный капитал», субсчет «Расчеты по вкладам в уставный капитал»
Дт 75 «Расчеты с учредителями», субсчет «Расчеты по выплате доходов» — Кт 68 «Расчеты по налогам и сборам»
Дт 84 «Нераспределенная прибыль (непокрытый убыток)» — Кт 75 «Расчеты с учредителями», субсчет «Расчеты по выплате доходов»
Дт 75 «Расчеты с учредителями», субсчет «Расчеты по выплате доходов» — Кт 75 «Расчеты с учредителями», субсчет «Расчеты по вкладам в уставной капитал»
Организация бухгалтерского учета на предприятии
Главная / Ответы на новые тесты / Организация бухгалтерского учета на предприятии / Страница 5 Упражнение 20395:Номер
Предъявленные и признанные штрафы за нарушение поставщиками условий хозяйственных договоров в бухгалтерском учете отражаются проводкой .
 ..
..Ответ:
 Дт 60 Кт 91 
 Дт 76 Кт 60 
 Дт 76 Кт 91 
Упражнение 20396:
Номер
При аккредитивной форме расчетов ...
Ответ:
 платежные документа оплачиваются по местонахождению поставщика через 10 дней после отгрузки им продукции 
 плательщик поручает обслуживающему его банку за счет своих депонированных средств или кредита оплатить товарно-материальные ценности (выполненные работы, оказанные услуги) по местонахождению получателя на условиях, предусмотренных договором 
 один аккредитив может быть использован для расчетов с несколькими поставщиками 
Упражнение 20397:
Номер
При вложении в уставный капитал другой организации передающая сторона приобретает .
 ..
..Ответ:
 акции этой организации 
 облигации 
 векселя 
Упражнение 20398:
Номер
При выбытии активов, принятых к бухгалтерскому учету в качестве финансовых вложений, по которым не определяется текущая рыночная стоимость, их стоимость организация определяет исходя из ...
Ответ:
 рыночной стоимости 
 стоимости по номиналу 
 последней оценки 
 средней первоначальной стоимости 
Упражнение 20399:
Номер
При наличии отклонения между фактической себестоимостью поступивших материалов и их стоимостью по учетным ценам составляется бухгалтерская запись .
 ..
..Ответ:
 Дт 15 Кт 60 
 Дт 10 Кт 15 
 Дт 10 Кт 16 
 Дт 16 Кт 15 
Упражнение 20400:
Номер
При открытии аккредитива за счет собственных средств дебетуют счет 55 «Специальные счета в банках» и кредитуют счет ...
Ответ:
 50 «Касса» 
 51 «Расчетные счета» 
 66 «Расчеты по краткосрочным кредитам и займам» 
Упражнение 20401:
Номер
При поступлении ТМЦ, оплаченных с аккредитива, в бухгалтерском учете покупателя делается запись .
 ..
..Ответ:
 Дт 10 Кт 60 
 Дт 10 Кт 51 
 Дт 60 Кт 51 
 Дт 10 Кт 55, субсчет «Аккредитивы» 
Упражнение 20402:
Номер
При признании в бухгалтерском учете выручки от продажи готовой продукции ее стоимость списывают со счета 43 «Готовая продукция» в дебет счетов ...
Ответ:
 90 «Продажи» 
 45 «Товары отгруженные» 
 91 «Прочие доходы и расходы» 
Упражнение 20403:
Номер
При продаже основных средств списание первоначальной стоимости отражается следующей бухгалтерской проводкой .
 ..
..Ответ:
 Дт 90 Кт 01 
 Дт 91 Кт 01, субсчет «Выбытие основных средств 
 Дт 91 Кт 08 
 Дт 01, субсчет «Выбытие основных средств» Кт 01 
Упражнение 20404:
Номер
При регистрации унитарного предприятия формирование уставного фонда отражается бухгалтерской записью ...
Ответ:
 Дт 80 Кт 75 
 Дт 75 Кт 80 
 Дт 79 Кт 80 
Упражнение 20405:
Номер
При создании резерва на оплату отпусков рабочих цехов основного производства делается запись .
 ..
..Ответ:
 Дт 96 Кт 20 
 Дт 20 Кт 96 
 Дт 70 Кт 96 
Упражнение 20406:
Номер
При страховом стаже работы до 5 лет пособие по временной нетрудоспособности оплачивается в размере ...
Ответ:
 80% 
 100% 
 60% 
 50% 
Упражнение 20407:
Номер
Прибыль направлена на выплату доходов учредителям и участникам
Ответ:
 Дт 99 Кт 75(70) 
 Дт 91 Кт 75(70) 
 Дт 84 Кт 75 (70) 
Упражнение 20408:
Номер
Прибыль от продажи продукции — это .
 ..
..Ответ:
 разница между фактической и плановой себестоимостью реализованной продукции 
 разница между выручкой от продажи продукции и ее полной фактической себестоимостью 
 выручка от продажи продукции 
Упражнение 20409:
Номер
Признаки, характеризующие полезность бухгалтерской информации согласно Концепции реформирования учета в РФ
Ответ:
 своевременность 
 прозрачность 
 ценность 
 уместность 
 надежность 
 сравнимость 
 правдивость 
Упражнение 20410:
Номер
Приняты к вычету по расчетам с бюджетом суммы НДС по полученным материалам
Ответ:
 Дт 68 Кт 19 
 Дт 10 Кт 68 
 Дт 68 Кт 91 
 Дт 68 Кт 90 
 Дт 91 Кт 68 
Упражнение 20411:
Номер
Приобретение оборудования, требующего монтажа отражается .
 ..
..Ответ:
 Дт 01 Кт 60, Дт 07 Кт 01 
 Дт 08 Кт 60, Дт 07 Кт 60 
 Дт 07 Кт 60, Дт 19 Кт 60 
Упражнение 20412:
Номер
Приходный и расходный кассовые ордера выписывает ...
Ответ:
 главный бухгалтер 
 кассир 
 руководитель организации 
Упражнение 20413:
Номер
Проводка операции перечисления средств транспортной организации за доставку материалов
Ответ:
 Дт 10 Кт 60 
 Дт 76 Кт 51 
 Дт 10 Кт 51 
Упражнение 20414:
Номер
Проводка, которой отражает поставщик предварительную оплату, полученную от покупателя
Ответ:
 Дт 51 Кт 60 
 Дт 51 Кт 62 
 Дт 60 Кт 51 
Упражнение 20415:
Номер
Проводка, которой отражают в учете начисление НДС
Ответ:
 Дт 68 Кт 90 
 Дт 90 Кт 68 
 Дт 68 Кт 43 
Упражнение 20416:
Номер
Проводка, которой отражают начисление амортизации НМА
Ответ:
 Дт 05 Кт 04 
 Дт 04 Кт 05 
 Дт 20 Кт 05 
 Дт 05 Кт 20 
Упражнение 20417:
Номер
Проводка, которой отражают начисление амортизации основных средств цеха
Ответ:
 Дт 01 Кт 02 
 Дт 25 Кт 02 
 Дт 02 Кт 25 
Упражнение 20418:
Номер
Проводка, которой отражают невозмещенные потери от брака
Ответ:
 Дт 20 Кт 28 
 Дт 28 Кт 20 
 Дт 43 Кт 28 
Упражнение 20419:
Номер
Проводка, которой отражают образование резерва для ремонта основных средств цеха
Ответ:
 Дт 25 Кт 96 
 Дт 96 Кт 25. 26 
26 
 Дт 96 Кт 23 
 Дт 01 Кт 96 
Упражнение 20420:
Номер
Проводка, которой отражают приобретение чековой книжки для расчетов чеками
Ответ:
 Дт 55/2 Кт 51 
 Дт 60 Кт 55/2 
 Дт 76 Кт 55/2 
Упражнение 20421:
Номер
Проводка, которой отражают списание себестоимости оказанных услуг, выполненных работ
Ответ:
 Дт 90 Кт 20 
 Дт 91 Кт 20 
 Дт 43 Кт 20 
Упражнение 20422:
Номер
Проводка, которой оформляется депонирование сумм заработной платы
Ответ:
 Дт 70 Кт 26 
 Дт 70 Кт 28 
 Дт 70 Кт 76/4 
 Дт 76/4 Кт 70 
Упражнение 20423:
Номер
Проводка, которой оформляется начисление пособия по временной нетрудоспособности
Ответ:
 Дт 69/1 Кт 70 
 Дт 68 Кт 70 
 Дт 70 Кт 69/1 
 Дт 20 Кт 70 
Упражнение 20424:
Номер
Проводка, которой оформляется удержание за брак
Ответ:
 Дт 68 Кт 70 
 Дт 69/2 Кт 70 
 Дт 28 Кт 70 
 Дт 70 Кт 28 
Упражнение 20425:
Номер
Проводка, которой оформляют операцию «акцептован счет поставщика за поступившие материалы на склад»
Ответ:
 Дт 10 Кт 60, Дт 19 Кт 60 
 Дт 10 Кт 60 
 Дт 10 Кт 62 
Упражнение 20426:
Номер
Проводка, которой оформляют отпуск материала со склада для упаковки готовой продукции на складе
Ответ:
 Дт 43 Кт 10 
 Дт 44 Кт 10 
 Дт 25 Кт 10 
Упражнение 20427:
Номер
Проводка, которой оформляют погашение недостачи кассиром
Ответ:
 Дт 94 Кт 50 
 Дт 50 Кт 94 
 Дт 50 Кт 73/2 
 Дт 73/2 Кт 50 
Упражнение 20428:
Номер
Проводка, которой оформляют реформацию баланса при получении прибыли по итогам года
Ответ:
 Дт 84 Кт 99 
 Дт 83 Кт 99 
 Дт 99 Кт 84 
Упражнение 20429:
Номер
Проводка, по которой заказчик отражает предварительную оплату услуг
Ответ:
 Дт 51 Кт 62 
 Дт 62 Кт 51 
 Дт 60 Кт 51 
Упражнение 20430:
Номер
Проводки, которыми отражают приобретение НМА у юридического лица
Ответ:
 Дт 08 Кт 76, Дт 19 Кт 76 
 Дт 04 Кт 08, Дт 04 Кт 19 
 Дт 04 Кт 76 
 Дт 04 Кт 60 
Упражнение 20431:
Номер
Проводки, которыми отражают приобретение у поставщика оборудования, не требующего монтажа
Ответ:
 Дт 01 Кт 60, Дт 19 Кт 60 
 Дт 08 Кт 60, Дт 19 Кт 60 
 Дт 19 Кт 60, Дт 01 Кт 08 
Упражнение 20432:
Номер
Проводки, которыми отражают списание НМА по истечении срока полезного использования
Ответ:
 Дт 05 Кт 04, Дт 91 Кт 04 
 Дт 91 Кт 04, Дт 99 Кт 04 
 Дт 04 Кт 91 
 Дт 05 Кт 04 
Упражнение 20433:
Номер
Проводки, котрыми отражают безвозмездное получение НМА
Ответ:
 Дт 04 Кт 98 
 Дт 08 Кт 98 
 Дт 04 Кт 08 
 Дт 04 Кт 83 
Упражнение 20434:
Номер
Проводки, отражающие предварительную оплату расходов по приобретению ценных бумаг
Ответ:
 Дт 58 Кт 51 
 Дт 51 Кт 58 
 Дт 76 Кт 51, Дт 58 Кт 76 
Упражнение 20435:
Номер
Произведены отчисления во внебюджетные фонды .
 ..
..Ответ:
 Дт 70 Кт 69 
 Дт 69 Кт 68 
 Дт 20 Кт 69 
 Дт 69 Кт 70 
Упражнение 20436:
Номер
Произведены отчисления от прибыли в резервный капитал
Ответ:
 Дт 86 Кт 99 
 Дт 91 Кт 86 
 Дт 99 Кт 86 
 Дт 84 Кт 82 
 Дт 91 Кт 83 
Упражнение 20437:
Номер
Пути привлечения заемных средств
Ответ:
 путем получения краткосрочных и долгосрочных займов от заимодавцев (кроме банков) внутри страны и за рубежом 
 путем продажи (выпуска) краткосрочных и долгосрочных облигаций, акций 
 путем финансовых векселей 
 привлечением кредитов и займов от юридических и физических лиц, а также за счет выпуска ценных бумаг 
Упражнение 20438:
Номер
Работнику основного производства с 20 сентября предоставлен ежегодный отпуск продолжительностью 28 календарных дней.
 Корреспонденция счетов по начислению отпускных
Корреспонденция счетов по начислению отпускныхОтвет:
 Дт 20 Кт 70 
 Дт 20 Кт 70, Дт 97 Кт 70 
 Дт 97 Кт 70 
Упражнение 20439:
Номер
Работы по монтажу оборудования считаются законченными если ...
Ответ:
 выполнен план капитальных вложений 
 составлены акты приемки выполненных работ 
 имеется в наличии акт об индивидуальной апробации оборудования 
Упражнение 20440:
Номер
Размер пособия по временной нетрудоспособности зависит от .
 ..
..Ответ:
 членства в профсоюзе 
 стажа работы 
 стажа работы и среднего заработка 
 среднего заработка 
Упражнение 20441:
Номер
Размер, в котором должен быть сформирован резервный капитал в акционерных обществах согласно действующему законодательству
Ответ:
 не менее 5% уставного капитала 
 не более 5% прибыли 
 25% уставного капитала 
 10% объема выручки 
Упражнение 20442:
Номер
Расход материалов, отпускаемых в производство и на другие нужды, ежедневно оформляют .
 ..
..Ответ:
 лимитно-заборными картами 
 карточками учета материалов 
 актами о приемке материалов 
Упражнение 20443:
Номер
Расходы будущих периодов — это ...
Ответ:
 средства, полученные в отчетном периоде, но относящиеся к будущим отчетным периодам 
 затраты, произведенные организацией в отчетном периоде 
 затраты, произведенные организацией в предшествующем и (или) отчетном периодах, но подлежащие включению в себестоимость продукции (работ, услуг) в последующие периоды 
Упражнение 20444:
Номер
Расходы на пользование кредитами банков, привлеченными на приобретение основных средств и нематериальных активов, после их ввода в эксплуатацию отражаются бухгалтерской записью .
 ..
..Ответ:
 Дт 26 Кт 66, 67 
 Дт 20 Кт 66, 67 
 Дт 91 Кт 66, 67 
 Дт 25 Кт 66, 67 
Технические характеристики — CloudTMS
Технические характеристики — CloudTMSZOOM TMS 101 — КАК НАЧАТЬ И СОЗДАТЬ УСПЕШНУЮ ПРАКТИКУ TMS Узнать больше
Этот сайт использует файлы cookie, чтобы помочь с функциональностью и анализировать использование вами наших услуг. Прочтите нашу Политику конфиденциальности.
Продолжать- 1. Описание
- 2. Использование по назначению
- 3. Рекомендуемое стандартное лечение
- 4. Катушки
- 5. Выходные параметры станка
- 6. Размеры и вес
1. Описание
Аппарат CloudTMS представляет собой систему повторяющейся транскраниальной магнитной стимуляции (rTMS). Это компьютеризированное медицинское устройство создает неинвазивные повторяющиеся импульсные магнитные поля достаточной величины для возбуждения нервных потенциалов в префронтальной коре для лечения большого депрессивного расстройства.
Принцип работы машины CloudTMS основан на разряде высоковольтного конденсатора (1,8 кВ) через стимулирующую катушку; Импульсное магнитное поле, создаваемое разрядным током (до 10 кА), проникает через нервно-мышечные ткани поблизости и индуцирует электрические токи в корковых нейронах.
Машина CloudTMS состоит из следующих основных компонентов:
- Основной блок магнитостимулятора
- Холодильный агрегат
- Дополнительный блок питания
- Охлаждаемый змеевик в форме восьмерки FEC-02-100-C
- Охлаждаемый змеевик в форме восьмерки AFEC-02-100-C
- Катушка в форме восьмерки FEC-02-100 (опция)
- Змеевик в форме восьмерки AFEC-02-100 (опция)
- Держатель катушки K8
- Гибкий рычаг K3 для позиционирования катушки
- Тележка с роликами
2.
 Использование по назначению
Использование по назначениюСистема CloudTMS показана для лечения большого депрессивного расстройства у взрослых пациентов, которым не удалось добиться удовлетворительного улучшения от предшествующего лечения антидепрессантами в текущем эпизоде.
3. Рекомендуемая стандартная обработка
| Напряженность магнитного поля | 120% Порог двигателя |
|---|---|
| Частота | 10 Гц |
| Продолжительность поезда | 4 секунды |
| Между поездами | 11 сек |
| Кол-во поездов | 75 |
| Магнитные импульсы за сеанс | 3000 |
| Продолжительность лечебного сеанса | ~ 19 мин. |
| Сессий в неделю | 5 |
| Схема лечения | 5 ежедневных занятий по 6 недель |
| Стимулируемая область мозга | Фронтальная кора |
4.
 Катушки
Катушки| Катушки (включая дополнительные аксессуары) | FEC-02-100-C, AFEC-02-100-C FEC-02-100 (дополнительно), AFEC-02-100 (дополнительно) |
|---|---|
| Конфигурация | Катушка в форме восьмерки |
| Материал сердечника | Воздушный сердечник |
| Охлаждение | FEC-02-100-C и AFEC-02-100- C: Жидкостное охлаждение FEC-02-100 и AFEC-02-100 Дополнительные аксессуары: Нет |
| Параметры катушки | FEC-02-100-C Внутренний диаметр — 47 × 50 мм1 Внешний диаметр — 97 × 100 мм1 Площадь = 184725 мм2 Средняя индуктивность 10 мкГн Плоская спиральная намотка N = 16 (4 витка / крыло x 2 крыла x 2 слоев) — AFEC-02-100-C Внутренний диаметр — 36 × 51 мм1 Внешний диаметр — 84 × 106 мм1 Площадь = 184725 мм2 Средняя индуктивность 10 мкГн Плоская спиральная намотка N = 16 (4 витка / крыло x 2 крылья x 2 слоя) — FEC-02-100 Внутренний диаметр — 47 × 50 мм1 Внешний диаметр — 97 × 100 мм1 Площадь = мм2 |
5.
 Выходные параметры станка
Выходные параметры станка| Амплитуда в единицах стандартного порога двигателя (SMT) | FEC-02-100-C 0 — 1,89 AFEC-02-100-C 0 — 2,38 FEC-02-100 0 — 1,92 AFEC-02-100 0 — 2,33 |
|---|---|
| Форма волны | Двухфазная синусоида |
| Активная длительность импульса (мкс) | 280 |
| Макс. Начальное значение дБ / dt (кТл / с) у поверхности катушки | FEC-02-100-C 25 AFEC-02-100-C 38 FEC-02-100 25 AFEC-02-100 32 |
| Система будет автоматически отключена, когда температура змеевика превысит: | 41 C (106 F) |
| Диапазон частот (Гц) | 0.1 — 30 (автономный) 0,1 — 100 (с ПК) |
| Диапазон длительности последовательности импульсов (с) | 0,5 — 100 |
| Диапазон интервалов между поездами (с) | 0–300 |
| Максимальное количество поездов за сеанс | 4800 = 2400 с [максимальная сессия] / (0,5 с [минимальная тренировка] + 0 с [минимальная пауза]) |
| Максимальное количество импульсов за сеанс | 72 000 (автономный режим) = 2400 с [макс. Сеанс] * 30 Гц 240 000 (с ПК) = 2400 с [макс. Сеанс] * 100 Гц Сеанс] * 30 Гц 240 000 (с ПК) = 2400 с [макс. Сеанс] * 100 Гц |
| Электробезопасность | Соответствует AAMI / ANSI ES 60601-1: 2005 / (R) 2012 и IEC 60601-1-2 |
| Соответствие стандартам ISO | Компания соответствует требованиям DIN EN ISO 13485: 2012 ISO 10993-1: 2009 ISO 14971: 2007 |
Рис.1 — Катушка магнитостимулятора
5. Размеры и масса станка
Каждый модуль CloudTMS имеет размеры W24 x D24 x h22, а размер тележки — W28 x D28 x h54.
Вес машины с тележкой составляет приблизительно 215 фунтов.
EpCAM: Структура и функция при здоровье и болезнях
Основные моменты
- •
Анализ структуры / функции EpCAM в (пато) физиологических условиях
- •
EpCAM также является мишенью для терапии и диагностики опухолей как регенеративная медицина
- •
Сигналы EpCAM в обогащенных тетраспанином микродоменах и посредством регулируемого протеолиза
- •
Эти модели возникают как общие механизмы передачи сигналов молекулами клеточной поверхности
294
- •
клеточная поверхность приводит к кишечным аномалиям
Abstract
Инъекция опухолевых клеток мышам более 30 лет назад привела к открытию эпителиального антигена, позже определенного как молекула клеточной адгезии (EpCAM). Хотя с тех пор EpCAM вызывал значительный интерес как мишень в терапии рака, механистические представления о функциях этого гликопротеина появились совсем недавно. Это могло быть вызвано множеством функций, приписываемых гликопротеину, его локализацией в различных субклеточных сайтах и сложными посттрансляционными модификациями. Здесь мы рассмотрим, как EpCAM изменяет силу межклеточной контактной адгезии и пластичность тканей, и как он регулирует пролиферацию и дифференцировку клеток.Будут выделены основные знания, полученные из болезней человека: мутантный EpCAM, который отсутствует на поверхности клетки, приводит к фатальным кишечным аномалиям (врожденная тафтинговая энтеропатия). EpCAM-опосредованная пролиферация клеток при раке может быть результатом передачи сигналов (i) посредством регулируемого внутримембранного протеолиза и / или (ii) локализации и ассоциации со связывающими партнерами в специализированных мембранных микродоменах. Новое понимание передачи сигналов EpCAM поможет разработать оптимизированные методы лечения рака и откроет новые возможности в области регенеративной медицины.
Хотя с тех пор EpCAM вызывал значительный интерес как мишень в терапии рака, механистические представления о функциях этого гликопротеина появились совсем недавно. Это могло быть вызвано множеством функций, приписываемых гликопротеину, его локализацией в различных субклеточных сайтах и сложными посттрансляционными модификациями. Здесь мы рассмотрим, как EpCAM изменяет силу межклеточной контактной адгезии и пластичность тканей, и как он регулирует пролиферацию и дифференцировку клеток.Будут выделены основные знания, полученные из болезней человека: мутантный EpCAM, который отсутствует на поверхности клетки, приводит к фатальным кишечным аномалиям (врожденная тафтинговая энтеропатия). EpCAM-опосредованная пролиферация клеток при раке может быть результатом передачи сигналов (i) посредством регулируемого внутримембранного протеолиза и / или (ii) локализации и ассоциации со связывающими партнерами в специализированных мембранных микродоменах. Новое понимание передачи сигналов EpCAM поможет разработать оптимизированные методы лечения рака и откроет новые возможности в области регенеративной медицины.
Ключевые слова
Молекула адгезии эпителиальных клеток (EpCAM)
Рак
Межклеточный контакт
Врожденная тафтинговая энтеропатия
Регулируемый внутримембранный протеолиз
Регулируемый внутримембранный протеолиз
Тетраспанин-обогащенный статьи © Elsevier BV, 2013. Опубликовано Elsevier BV. Все права защищены.
Рекомендуемые статьи
Цитирующие статьи
% PDF-1.5
%
4 0 obj
>
эндобдж
7 0 объект
(\ 376 \ 377 \ 000I \ 000n \ 000t \ 000r \ 000o \ 000d \ 000u \ 000c \ 000t \ 000i \ 000o \ 000n \ 000 \ 040 \ 000t \ 000o \ 000 \ 040 \ 000S \ 000i \ 000m \ 000u \ 000л \ 000i \ 000n \ 000k)
эндобдж
8 0 объект
>
эндобдж
11 0 объект
(\ 376 \ 377 \ 000S \ 000o \ 000l \ 000v \ 000i \ 000n \ 000g \ 000 \ 040 \ 000a \ 000n \ 000 \ 040 \ 000O \ 000D \ 000E)
эндобдж
12 0 объект
>
эндобдж
15 0 объект
(\ 376 \ 377 \ 000H \ 000a \ 000n \ 000d \ 000l \ 000i \ 000n \ 000g \ 000 \ 040 \ 000T \ 000i \ 000m \ 000e \ 000 \ 040 \ 000i \ 000n \ 000 \ 040 \ 000F \ 000i \ 000r \ 000s \ 000t \ 000 \ 040 \ 000O \ 000r \ 000d \ 000e \ 000r \ 000 \ 040 \ 000D \ 000i \ 000f \ 000f \ 000e \ 000r \ 000e \ 000n \ 000t \ 000i \ 000a \ 000l \ 000 \ 040 \ 000E \ 000q \ 000u \ 000a \ 000t \ 000i \ 000o \ 000n \ 000s)
эндобдж
16 0 объект
>
эндобдж
19 0 объект
(\ 376 \ 377 \ 000W \ 000o \ 000r \ 000k \ 000i \ 000n \ 000g \ 000 \ 040 \ 000w \ 000i \ 000t \ 000h \ 000 \ 040 \ 000S \ 000i \ 000m \ 000u \ 000l \ 000i \ 000n \ 000k \ 000 \ 040 \ 000O \ 000u \ 000t \ 000p \ 000u \ 000t)
эндобдж
20 0 объект
>
эндобдж
23 0 объект
(\ 376 \ 377 \ 000P \ 000r \ 000i \ 000n \ 000t \ 000i \ 000n \ 000g \ 000 \ 040 \ 000S \ 000i \ 000m \ 000u \ 000l \ 000i \ 000n \ 000k \ 000 \ 040 \ 000S \ 000c \ 000o \ 000p \ 000e \ 000 \ 040 \ 000I \ 000m \ 000a \ 000g \ 000e \ 000s)
эндобдж
24 0 объект
>
эндобдж
27 0 объект
(\ 376 \ 377 \ 000S \ 000c \ 000i \ 000l \ 000a \ 000b \ 000 \ 040 \ 000a \ 000n \ 000d \ 000 \ 040 \ 000X \ 000c \ 000o \ 000s)
эндобдж
28 0 объект
>
эндобдж
31 0 объект
(\ 376 \ 377 \ 000F \ 000i \ 000r \ 000s \ 000t \ 000 \ 040 \ 000O \ 000r \ 000d \ 000e \ 000r \ 000 \ 040 \ 000O \ 000D \ 000E \ 000s \ 000 \ 040 \ 000i \ 000n \ 000 \ 040 \ 000M \ 000A \ 000T \ 000L \ 000A \ 000B)
эндобдж
32 0 объект
>
эндобдж
35 0 объект
(\ 376 \ 377 \ 000E \ 000x \ 000e \ 000r \ 000c \ 000i \ 000s \ 000e \ 000s)
эндобдж
36 0 объект
>
эндобдж
39 0 объект
(\ 376 \ 377 \ 000F \ 000i \ 000r \ 000s \ 000t \ 000 \ 040 \ 000O \ 000r \ 000d \ 000e \ 000r \ 000 \ 040 \ 000D \ 000i \ 000f \ 000f \ 000e \ 000r \ 000e \ 000n \ 000t \ 000i \ 000a \ 000l \ 000 \ 040 \ 000E \ 000q \ 000u \ 000a \ 000t \ 000i \ 000o \ 000n \ 000s)
эндобдж
40 0 объект
>
эндобдж
43 0 объект
(\ 376 \ 377 \ 000E \ 000x \ 000p \ 000o \ 000n \ 000e \ 000n \ 000t \ 000i \ 000a \ 000l \ 000 \ 040 \ 000G \ 000r \ 000o \ 000w \ 000t \ 000h \ 000 \ 040 \ 000a \ 000n \ 000d \ 000 \ 040 \ 000D \ 000e \ 000c \ 000a \ 000y)
эндобдж
44 0 объект
>
эндобдж
47 0 объект
(\ 376 \ 377 \ 000N \ 000e \ 000w \ 000t \ 000o \ 000n \ 000 ‘\ 000s \ 000 \ 040 \ 000L \ 000a \ 000w \ 000 \ 040 \ 000o \ 000f \ 000 \ 040 \ 000C \ 000o \ 000o \ 000l \ 000i \ 000n \ 000g)
эндобдж
48 0 объект
>
эндобдж
51 0 объект
(\ 376 \ 377 \ 000F \ 000r \ 000e \ 000e \ 000 \ 040 \ 000F \ 000a \ 000l \ 000l \ 000 \ 040 \ 000w \ 000i \ 000t \ 000h \ 000 \ 040 \ 000D \ 000r \ 000a \ 000g)
эндобдж
52 0 объект
>
эндобдж
55 0 объект
(\ 376 \ 377 \ 000P \ 000u \ 000r \ 000s \ 000u \ 000i \ 000t \ 000 \ 040 \ 000C \ 000u \ 000r \ 000v \ 000e \ 000s)
эндобдж
56 0 объект
>
эндобдж
59 0 объект
(\ 376 \ 377 \ 000T \ 000h \ 000e \ 000 \ 040 \ 000L \ 000o \ 000g \ 000i \ 000s \ 000t \ 000i \ 000c \ 000 \ 040 \ 000E \ 000q \ 000u \ 000a \ 000t \ 000i \ 000o \ 000n)
эндобдж
60 0 объект
>
эндобдж
63 0 объект
(\ 376 \ 377 \ 000T \ 000h \ 000e \ 000 \ 040 \ 000L \ 000o \ 000g \ 000i \ 000s \ 000t \ 000i \ 000c \ 000 \ 040 \ 000E \ 000q \ 000u \ 000a \ 000t \ 000i \ 000o \ 000n \ 000 \ 040 \ 000w \ 000i \ 000t \ 000h \ 000 \ 040 \ 000D \ 000e \ 000l \ 000a \ 000y)
эндобдж
64 0 объект
>
эндобдж
67 0 объект
(\ 376 \ 377 \ 000E \ 000x \ 000e \ 000r \ 000c \ 000i \ 000s \ 000e \ 000s)
эндобдж
68 0 объект
>
эндобдж
71 0 объект
(\ 376 \ 377 \ 000S \ 000e \ 000c \ 000o \ 000n \ 000d \ 000 \ 040 \ 000O \ 000r \ 000d \ 000e \ 000r \ 000 \ 040 \ 000D \ 000i \ 000f \ 000f \ 000e \ 000r \ 000e \ 000n \ 000t \ 000i \ 000a \ 000l \ 000 \ 040 \ 000E \ 000q \ 000u \ 000a \ 000t \ 000i \ 000o \ 000n \ 000s)
эндобдж
72 0 объект
>
эндобдж
75 0 объект
(\ 376 \ 377 \ 000C \ 000o \ 000n \ 000s \ 000t \ 000a \ 000n \ 000t \ 000 \ 040 \ 000C \ 000o \ 000e \ 000f \ 000f \ 000i \ 000c \ 000i \ 000e \ 000n \ 000t \ 000 \ 040 \ 000E \ 000q \ 000u \ 000a \ 000t \ 000i \ 000o \ 000n \ 000s)
эндобдж
76 0 объект
>
эндобдж
79 0 объект
(\ 376 \ 377 \ 000P \ 000r \ 000o \ 000j \ 000e \ 000c \ 000t \ 000i \ 000l \ 000e \ 000 \ 040 \ 000M \ 000o \ 000t \ 000i \ 000o \ 000n)
эндобдж
80 0 объект
>
эндобдж
83 0 объект
(\ 376 \ 377 \ 000T \ 000h \ 000e \ 000 \ 040 \ 000B \ 000o \ 000u \ 000n \ 000c \ 000i \ 000n \ 000g \ 000 \ 040 \ 000B \ 000a \ 000l \ 000l)
эндобдж
84 0 объект
>
эндобдж
87 0 объект
(\ 376 \ 377 \ 000N \ 000o \ 000n \ 000l \ 000i \ 000n \ 000e \ 000a \ 000r \ 000 \ 040 \ 000P \ 000e \ 000n \ 000d \ 000u \ 000l \ 000u \ 000m \ 000 \ 040 \ 000A \ 000n \ 000i \ 000m \ 000a \ 000t \ 000i \ 000o \ 000n)
эндобдж
88 0 объект
>
эндобдж
91 0 объект
(\ 376 \ 377 \ 000S \ 000e \ 000c \ 000o \ 000n \ 000d \ 000 \ 040 \ 000O \ 000r \ 000d \ 000e \ 000r \ 000 \ 040 \ 000O \ 000D \ 000E \ 000s \ 000 \ 040 \ 000i \ 000n \ 000 \ 040 \ 000M \ 000A \ 000T \ 000L \ 000A \ 000B)
эндобдж
92 0 объект
>
эндобдж
95 0 объект
(\ 376 \ 377 \ 000E \ 000x \ 000e \ 000r \ 000c \ 000i \ 000s \ 000e \ 000s)
эндобдж
96 0 объект
>
эндобдж
99 0 объект
(\ 376 \ 377 \ 000T \ 000r \ 000a \ 000n \ 000s \ 000f \ 000e \ 000r \ 000 \ 040 \ 000F \ 000u \ 000n \ 000c \ 000t \ 000i \ 000o \ 000n \ 000s \ 000 \ 040 \ 000a \ 000n \ 000d \ 000 \ 040 \ 000S \ 000t \ 000a \ 000t \ 000e \ 000 \ 040 \ 000S \ 000p \ 000a \ 000c \ 000e \ 000 \ 040 \ 000B \ 000l \ 000o \ 000c \ 000k \ 000s)
эндобдж
100 0 объект
>
эндобдж
103 0 объект
(\ 376 \ 377 \ 000S \ 000t \ 000a \ 000t \ 000e \ 000 \ 040 \ 000S \ 000p \ 000a \ 000c \ 000e \ 000 \ 040 \ 000F \ 000o \ 000r \ 000m \ 000u \ 000l \ 000a \ 000t \ 000i \ 000o \ 000n)
эндобдж
104 0 объект
>
эндобдж
107 0 объект
(\ 376 \ 377 \ 000T \ 000r \ 000a \ 000n \ 000s \ 000f \ 000e \ 000r \ 000 \ 040 \ 000F \ 000u \ 000n \ 000c \ 000t \ 000i \ 000o \ 000n \ 000s)
эндобдж
108 0 объект
>
эндобдж
111 0 объект
(\ 376 \ 377 \ 000S \ 000y \ 000s \ 000t \ 000e \ 000m \ 000s \ 000 \ 040 \ 000o \ 000f \ 000 \ 040 \ 000D \ 000i \ 000f \ 000f \ 000e \ 000r \ 000e \ 000n \ 000t \ 000i \ 000a \ 000l \ 000 \ 040 \ 000E \ 000q \ 000u \ 000a \ 000t \ 000i \ 000o \ 000n \ 000s)
эндобдж
112 0 объект
>
эндобдж
115 0 объект
(\ 376 \ 377 \ 000L \ 000i \ 000n \ 000e \ 000a \ 000r \ 000 \ 040 \ 000S \ 000y \ 000s \ 000t \ 000e \ 000m \ 000s)
эндобдж
116 0 объект
>
эндобдж
119 0 объект
(\ 376 \ 377 \ 000N \ 000o \ 000n \ 000l \ 000i \ 000n \ 000e \ 000a \ 000r \ 000 \ 040 \ 000M \ 000o \ 000d \ 000e \ 000l \ 000s)
эндобдж
120 0 объект
>
эндобдж
123 0 объект
(\ 376 \ 377 \ 000I \ 000n \ 000d \ 000e \ 000x)
эндобдж
124 0 объект
>
эндобдж
128 0 obj>
поток
x ڍ RMo0WJKk7 &
! FA6m’4 _- «P ٱ_ [h2y
2Π= 4l45
ԡ + p # / x / ^ K, DɍGj. SFvg y,) SXLVVVͤjr; | i۶mm۶A W6 $ » O ٳ Ύ; & ח / _ „TY.] * ˢE & ~ BAèw6oΝ; gelxbbb Gihh
SFvg y,) SXLVVVͤjr; | i۶mm۶A W6 $ » O ٳ Ύ; & ח / _ „TY.] * ˢE & ~ BAèw6oΝ; gelxbbb Gihh
Коэнзим Q пептоид, нацеленный на митохондрии, вызывает стресс в растительных клетках и устраняет супероксидный салин.
FAO. Как накормить мир в 2050 году. С. 35 (2009).
Флауэрс, Дж. Т. Повышение солеустойчивости сельскохозяйственных культур. J. Exp. Бот. 55 (396), 307–319 (2004).
CAS Google Scholar
Маннс Р. и Тестер М. Механизмы солености. Annu. Rev. Plant Biol. 59 , 651–681 (2008).
CAS Google Scholar
Zhu, B.-Z., Zhao, H.-T., Kalyanaraman, B. & Frei, B. Металл-независимое образование гидроксильных радикалов галогенированными хинонами и перекисью водорода: исследование спинового захвата ESR . Free Radic. Биол. Med. 32 (5), 465–473 (2002).
CAS Google Scholar

Маннс, Р. Гены и солеустойчивость: объединяя их. New Phytol. 167 , 645–663 (2005).
CAS Google Scholar
Hazman, M., Hause, B., Eiche, E., Riemann, M. и Nick, P. Различные формы осмотического стресса вызывают качественно разные реакции у риса. J. Plant Physiol. 202 , 45–56 (2016).
CAS Google Scholar
Кацухара М. и Шибасака М. Смерть клеток и восстановление роста ячменя после кратковременного солевого стресса. J. Plant Res. 113 , 239–243 (2000).
CAS Google Scholar
Ismail, A. et al. Адаптация к соли требует эффективной тонкой настройки передачи сигналов жасмоната. Protoplasma 251 , 881–898 (2014).
CAS Google Scholar
Миттлер Р. Окислительный стресс, антиоксиданты и стрессоустойчивость. Trends Plant Sci. 7 (9), 405–410 (2002).
Окислительный стресс, антиоксиданты и стрессоустойчивость. Trends Plant Sci. 7 (9), 405–410 (2002).
CAS Google Scholar
Апель К. и Хирт Х. Активный кислород: метаболизм, окислительный стресс и передача сигналов. Annu. Rev. Plant Biol. 55 , 373–399 (2004).
CAS Google Scholar
Фойер, Х. К. и Ноктор, Г. Редокс-гомеостаз и передача сигналов антиоксидантами: метаболический интерфейс между восприятием стресса и физиологическими реакциями. Растительная клетка 17 , 1866–1875 (2005).
CAS PubMed Central PubMed Google Scholar
Zhu, J.-K. Сигналы и ответы на абиотический стресс у растений. Ячейка 167 , 313–324 (2016).
CAS PubMed Central PubMed Google Scholar
Миллер Г., Сузуки Н., Чифтчи-Йилмаз С. и Миттлер Р. Гомеостаз и сигнализация активных форм кислорода во время стрессов засухи и засоления. Plant Cell Environ. 33 , 453–467 (2010).
CAS Google Scholar
Марино Д., Дананд К., Пуппо А. и Паули Н. Всплеск НАДФН-оксидаз в растениях. Trends Plant Sci. 17 (1), 9–15 (2012).
CAS Google Scholar
Price, H.A. et al. Окислительные сигналы в табаке повышают содержание кальция в цитозоле. Растительная клетка 6 , 1301–1310 (1994).
ADS CAS PubMed Central PubMed Google Scholar
Testerink, C. & Munnik, T. Молекулярные, клеточные и физиологические реакции на образование фосфатидной кислоты в растениях. J. Exp. Бот. 62 (7), 2349–2361 (2011).
CAS Google Scholar

Нюрнбергер, Т. Восприятие сигналов в защите растений от патогенов. CMLS Cell Mol. Life Sci. 55 , 167–182 (1999).
Google Scholar
Adachi, H. et al. Факторы транскрипции WRKY, фосфорилированные MAPK Регулируют иммунную НАДФН-оксидазу растений в Nicotiana benthamiana . Растительная клетка 27 , 2645–2663 (2015).
CAS PubMed Central PubMed Google Scholar
Zancani, M. et al. Переход проницаемости в митохондриях растений: недостающее звено. Перед. Plant Sci. 6 (1120), 1–8 (2015).
Google Scholar
Ng, S. et al. Циклинзависимая киназа E1 (CDKE1) обеспечивает клеточный переключатель у растений между ответами роста и стресса. J. Biol. Chem. 288 (5), 3449–3459 (2013).
CAS Google Scholar
.bmp)
De Clercq, I. et al. Связанный с мембраной фактор транскрипции NAC ANAC013 участвует в митохондриальной ретроградной регуляции реакции на окислительный стресс у Arabidopsis. Растительная клетка 25 , 3472–3490 (2013).
PubMed Central PubMed Google Scholar
Ng, S. et al. Связанный с мембраной фактор транскрипции NAC, ANAC017, опосредует митохондриальную ретроградную передачу сигналов у Arabidopsis. Растительная клетка 25 , 3450–3471 (2013).
CAS PubMed Central PubMed Google Scholar
Каруппанапандиан, Т., Мун, Дж.-К., Ким, К., Манохаран, К. и Ким, В. Активные формы кислорода в растениях: их генерация, передача сигнала и механизмы поглощения. AJCS 5 (6), 709–725 (2011).
CAS Google Scholar
Роллан, А. Доставка ядерных генов: подход троянского коня. Мнение эксперта. Препарат Делив. 3 (1), 1–10 (2006).
Доставка ядерных генов: подход троянского коня. Мнение эксперта. Препарат Делив. 3 (1), 1–10 (2006).
CAS Google Scholar
Вагстафф, К. М. и Янс, Д. А. Трансдукция белка: проникающие в клетки пептиды и их терапевтическое применение. Curr. Med. Chem. 13 , 1371–1387 (2006).
CAS Google Scholar
Järver, P. & Langel, Ü. Проникающие в клетки пептиды — краткое введение. Biochem. Биофиз. Acta. 1758 , 260–263 (2006).
Google Scholar
Торчилин В.П. Внутриклеточная доставка фармацевтических наноносителей, опосредованная Tatp. Biochem. Soc. Пер. 35 (4), 816–820 (2007).
CAS Google Scholar
Чу, А., Амундсен, Э. и Эдес, Ф. Транслокация проникающих в клетки пептидов и доставка их грузов в микроспорах тритикале. Plant Cell Rep. 28 , 801–810 (2009).
Plant Cell Rep. 28 , 801–810 (2009).
CAS Google Scholar
Mäe, M. et al. Интернализация проникающих в клетки пептидов в протопласты табака. Biochem. Биофиз. Acta. 1669 , 101–107 (2005).
Google Scholar
Chugh, A. & Eudes, F. Изучение поглощения проникающих в клетки пептидов и их грузов незрелыми зародышами пшеницы с проницаемостью. FEBS J. 275 , 2403–2414 (2008).
CAS Google Scholar
Эггенбергер, К., Бирталан, Э., Шредер, Т., Брезе, С. и Ник, П. Прохождение троянских пептоидов в клетки растений. ChemBioChem 10 , 2504–2512 (2009).
CAS Google Scholar
Simon, R.J. et al. Пептоиды: модульный подход к открытию лекарств. Proc. Natl. Акад. Sci. США 89 , 9367–9371 (1992).
Natl. Акад. Sci. США 89 , 9367–9371 (1992).
ADS CAS Google Scholar
Olivos, H.J., Alluri, P.G., Reddy, M.M., Salony, D. & Kodadek, T. Твердофазный синтез пептоидов с помощью микроволнового излучения. Org. Lett. 4 (23), 4057–4059 (2002).
CAS Google Scholar
Miller, M. S. et al. Сравнение протеолитической восприимчивости гомологичных 1-аминокислоты, d-аминокислоты и N-замещенного пептида глицина и пептоидных олигомеров. Drug Dev. Res. 35 , 20–32 (1995).
ADS CAS Google Scholar
Сот, Дж. М. и Новик, С. Дж. Неестественные олигомеры и библиотеки неприродных олигомеров. Curr. Opin. Chem. Биол. 1 , 120–129 (1997).
CAS Google Scholar
Asfaw, K. G. et al. Пептоид доставляет производное CoQ в митохондрии растений посредством эндоцитоза. Sci. Реп 9 , 1–18 (2019).
Google Scholar
Хуанг, X., Майш, Дж. И Ник, П. Сенсорная роль актина в ауксин-зависимых ответах табака BY-2. J. Plant Physiol. 218 , 6–15 (2017).
CAS Google Scholar
Мольц, Ф. Дж. Водные потенциалы, индуцированные ростом в клетках и тканях растений. Plant Physiol. 62 , 423–429 (1978).
CAS PubMed Central PubMed Google Scholar
Гленн, Э. П. и Браун, Дж. Дж. Солеустойчивость и потенциал сельскохозяйственных культур галофитов. Crit. Rev. Plant Sci. 18 (2), 227–255 (1999).
Google Scholar
Хит, Р. Л. и Пакер, Л. Фотопероксидация в изолированных хлоропластах: I. Кинетика и стехиометрия перекисного окисления жирных кислот. Arch. Биохим. Биофиз. 125 , 189–198 (1968).
и Пакер, Л. Фотопероксидация в изолированных хлоропластах: I. Кинетика и стехиометрия перекисного окисления жирных кислот. Arch. Биохим. Биофиз. 125 , 189–198 (1968).
CAS Google Scholar
Исмаил А., Такеда С. и Ник П. Жизнь и смерть в условиях солевого стресса: одни и те же игроки, разное время ?. J. Exp. Бот. 65 (12), 2963–2979 (2014).
CAS Google Scholar
Гулд, С. Б., Валлер, Р. Ф. и Макфадден, Г. И. Эволюция пластидов. Annu. Rev. Plant Biol. 59 , 491–517 (2008).
CAS Google Scholar
Blanco, N.E., Guinea-Dıaz, M., Whelan, J. & Strand, A. Взаимодействие между пластидными и митохондриальными ретроградными сигнальными путями во время изменений окислительно-восстановительного статуса пластид. Philos. Пер. R. Soc. B 369 , 1–8 (2014).
Google Scholar
Аллен, Дж. Ф. Почему хлоропласты и митохондрии сохраняют свои собственные геномы и генетические системы: колокация для окислительно-восстановительной регуляции экспрессии генов. PNAS 112 (33), 10231–10238 (2015).
ADS CAS Google Scholar
Hartl, M. & Finkemeier, I.В стадию вступают посттрансляционные модификации ретроградной передачи сигналов митохондрий растений. Перед. Plant Sci. 3 (253), 1–7 (2012).
Google Scholar
Börner, T. Открытие ретроградной передачи сигналов от пластида к ядру — личная перспектива. Protoplasma 254 , 1845–1855 (2017).
PubMed Central PubMed Google Scholar
Кляйне Т. и Дарио Лейстер Д. Ретроградная передача сигналов: органеллы объединяются. Biochim. Биофиз. Acta 1857 , 1313–1325 (2016).
Biochim. Биофиз. Acta 1857 , 1313–1325 (2016).
CAS Google Scholar
Ho, L.H. M. et al. Идентификация регуляторных путей, контролирующих экспрессию генов стресс-чувствительных митохондриальных белков у Arabidopsis. Plant Physiol. 147 , 858–1873 (2008).
Google Scholar
Ng, S. et al. Антероградная и ретроградная регуляция ядерных генов, кодирующих митохондриальные белки во время роста, развития и стресса. Мол. Завод 7 , 1075–1093 (2014).
CAS Google Scholar
Хоанг, X. Л. Т., Нхи, Д. Н. Х., Тху, Н. Б. А., Тао, Н. П. и Тран, Л.-С.П. Факторы транскрипции и их роль в передаче сигнала у растений в условиях абиотических стрессов. Curr. Геномика 18 , 483–497 (2017).
PubMed Central PubMed Google Scholar

Gestel, K. V., Köhler, R.H., Verbelen, J.-P. Митохондрии растений перемещаются на F-актине, но их положение в кортикальной цитоплазме зависит как от F-актина, так и от микротрубочек. J. Exp. Бот. 53 (369), 659–667 (2002).
Google Scholar
Флис, В. В. и Даум, Г. Транспорт липидов между эндоплазматическим ретикулумом и митохондриями. Колд Спринг Харб. Перспектива. Биол. 5 (6), 1-22 (2013).
Google Scholar
Моллер М. И. Что горячего в митохондриях растений ?. Physiol. Растение. 157 , 256–263 (2016).
CAS Google Scholar
Хуанг, С., Акен, В. О., Шварцлендер, М., Белт, К., Миллар, Х. А. Роль митохондриальных активных форм кислорода в клеточной передаче сигналов и стрессовой реакции у растений. Plant Physiol. 171 , 1551–1559 (2016).
CAS PubMed Central PubMed Google Scholar
Вагнер, С., Акен, О. В., Эльзассер, М. и Шварцлендер, М. Передача сигналов митохондриальной энергии и ее роль в реакции растений на стресс с низким содержанием кислорода. Plant Physiol. 176 , 1156–1170 (2018).
CAS PubMed Central PubMed Google Scholar
Gulaboski, R. et al. Гидроксилированные производные диметокси-1,4-бензохинона в качестве окислительно-восстановительных лигандов щелочноземельных металлов и акцепторов радикалов. Sci. Отчет 3 (1865), 1–8 (2013).
Google Scholar
Candas, D. & Li, J. J. MnSOD в регуляции потенциала реакции на окислительный стресс через приток митохондриального белка. Антиоксид. Редокс-сигнал. 20 (10), 1599–1617 (2014).
CAS PubMed Central PubMed Google Scholar
Li, C.-R. et al. Раскрытие ретроградной регуляции митохондрий при индукции абиотического стресса у риса АЛЬТЕРНАТИВНАЯ ОКСИДАЗА1 гены. Plant Cell Environ. 36 , 775–788 (2013).
ADS CAS Google Scholar
Møller, I. M. & Sweetlove, L. J. Передача сигналов ROS — требуется специфичность. Trends Plant Sci. 15 , 370–374 (2010).
Google Scholar
Holzmeister, C. et al. Дифференциальное ингибирование супероксиддисмутаз Arabidopsis путем нитрования тирозина, опосредованного пероксинитритом. J. Exp. Бот. 66 (3), 989–999 (2015).
CAS Google Scholar
Sunkar, R., Kapoor, A. & Zhu, J.-K. Посттранскрипционная индукция двух генов супероксиддисмутазы Cu / Zn у арабидопсиса опосредуется подавлением miR398 и важна для устойчивости к окислительному стрессу. Растительная клетка 18 , 2051–2065 (2006).
Растительная клетка 18 , 2051–2065 (2006).
CAS PubMed Central PubMed Google Scholar
Лук, Э., Янг, М., Дженсен, Л. Т., Бурбонне, Ю. и Кюло, В. С. Активация супероксиддисмутазы 2 марганцем в митохондриях Saccharomyces cerevisiae . J. Biol. Chem. 280 (24), 22715–22720 (2005).
CAS Google Scholar
Кулотта В. К., Янг М. и О’Халлоранб Т. В. Активация супероксиддисмутаз: прижимание металла к педали. Biochim. Биофиз.Acta 1763 (7), 747–758 (2006).
CAS PubMed Central PubMed Google Scholar
Ismail, A., Seo, M., Takebayashi, Y., Kamiya, Y. & Nick, P. Сбалансированный статус JA / ABA может коррелировать с адаптацией к осмотическому стрессу в клетках Vitis. J. Plant Physiol. 185 , 57–64 (2015).
CAS Google Scholar
Исмаил А., Риманн М. и Ник П. Жасмонатный путь опосредует солеустойчивость виноградных лоз. J. Exp. Бот. 63 (5), 2127–2139 (2012).
CAS PubMed Central PubMed Google Scholar
Akaberi, S. et al. Гидропероксидлиаза жирных кислот виноградной лозы вырабатывает летучие вещества, разрушающие актин, и способствует гибели клеток, связанной с защитой. J. Exp. Бот. 69 (12), 2883–2896 (2018).
CAS PubMed Central PubMed Google Scholar
Park, S. W. et al. Циклофилин 20–3 передает сигнал 12-оксофитодиеновой кислоты во время чувствительной к стрессу регуляции клеточного окислительно-восстановительного гомеостаза. Proc. Natl. Акад. Sci. 110 (23), 9559–9564 (2013).
ADS CAS Google Scholar
Estavillo, G.M. et al. Доказательства ретроградного пути хлоропластов SAL1-PAP, который функционирует в условиях засухи и высокой световой сигнализации у Arabidopsis. Растительная клетка 23 , 3992-4012 (2011).
CAS PubMed Central PubMed Google Scholar
Jaipargas, E.-A., Mathur, N., Daher, F. B., Wasteneys, G.O. & Mathur, J. Высокая интенсивность света приводит к усилению взаимодействия пероксул и митохондрий у растений. Перед. Cell Dev. Биол. 4 (6), 1–11 (2016).
Google Scholar
Bueno, P., Varela, J., Cimenez-Gallego, C. & del Rio, L.A. Пероксисомальная медь, супероксиддисмутаза цинка: характеристика изофермента семядолей арбуза. Plant Physiol. 108 , 1151–1160 (1995).
CAS PubMed Central PubMed Google Scholar

дель Рио, Л. А., Сандалио, Л. М., Алтомаре, Д. А. и Зилинскас, Б. А. Митохондриальная и пероксисомальная супероксиддисмутаза марганца: дифференциальная экспрессия во время старения листьев. J. Exp. Бот. 54 (384), 923–933 (2003).
Google Scholar
Hamilton, A.J., Holdom, M.D. и Jeavons, L. Экспрессия Cu, Zn-супероксиддисмутазы Aspergillus fumigatu s, как определено с помощью иммунохимии и иммуноэлектронной микроскопии. FEMS Immunol. Med. Microbiol. 14 , 95–102 (1996).
CAS Google Scholar
Smith, C.A., Melino, V.J., Sweetman, C. & Soole, K. L. Манипуляции с альтернативной оксидазой могут влиять на солевую толерантность Arabidopsis thaliana . Physiol. Растение. 137 , 459–472 (2009).
CAS Google Scholar
Jardim-Messeder, D. et al. Сукцинатдегидрогеназа (митохондриальный комплекс II) является источником активных форм кислорода в растениях и регулирует развитие и реакцию на стресс. New Phytol. 208 , 776–789 (2015).
et al. Сукцинатдегидрогеназа (митохондриальный комплекс II) является источником активных форм кислорода в растениях и регулирует развитие и реакцию на стресс. New Phytol. 208 , 776–789 (2015).
CAS Google Scholar
Нагата Т., Немото Ю. и Хасезо С. Клеточная линия табака BY-2 в качестве клетки «HeLa» в клеточной биологии высших растений. Внутр. Rev. Cytol. 132 , 1–30 (1992).
CAS Google Scholar
Абубакар Л.А. Влияние засоления на параметры роста галотолерантных микроводорослей, Dunaliella spp . Нигер. J. Basic Appl. Sci. 24 (2), 85–91 (2016).
Google Scholar
Ник П., Хойинг А. и Эманн Б. Шаперонины растений: роль в формировании стенок, зависимых от микротрубочек ?. Protoplasma 211 , 234–244 (2000).
CAS Google Scholar
Гафф, Ф. Д. и Оконг’О-Огола, О. Использование непроникающих пигментов для тестирования выживаемости клеток. J. Exp. Бот. 22 , 756–758 (1971).
Google Scholar
Хендерсон, М. Л. и Чаппелл, Б. Дж. Дигидрородамин 123: флуоресцентный зонд для образования супероксида ?. Eur. J. Biochem. 217 , 973–980 (1993).
CAS Google Scholar
Ходжсон, Дж. А. Р. и Рейсон, К. Дж. Перекисное окисление липидов и активность супероксиддисмутазы в отношении фотоингибирования, вызванного охлаждением при умеренном свете. Planta 185 , 215–219 (1991).
CAS Google Scholar
Lata, C., Jha, S., Dixit, V., Sreenivasulu, N. & Prasad, M. Дифференциальные антиоксидантные реакции на вызванный обезвоживанием окислительный стресс у основных сортов проса лисохвоста [ Setaria italica (L.)]. Protoplasma 248 , 817–828 (2011).
CAS Google Scholar
Vacca, A. R. et al. Производство активных форм кислорода, изменение цитозольной аскорбатпероксидазы и нарушение митохондриального метаболизма являются ранними событиями в вызванной тепловым шоком запрограммированной гибели клеток в клетках Tobacco Bright-Yellow 2. Plant Physiol. 134 , 1100–1112 (2004).
CAS PubMed Central PubMed Google Scholar
де Пинто, К. М., Парадизо, А., Леонетти, П. и де Гара, Л. Пероксид водорода, оксид азота и цитозольная аскорбатпероксидаза на перекрестке между защитой и гибелью клеток. Плант Дж. 48 , 784–795 (2006).
Google Scholar
Beauchamp, C. & Fridovich, I. Супероксиддисмутаза: улучшенные анализы и анализ, применимый к акриламидным гелям. Анал. Биохим. 44 , 276–287 (1971).
CAS Google Scholar
Weydert, J. C. & Cullen, J. J. Измерение супероксиддисмутазы, каталазы и глутатионпероксидазы в культивируемых клетках и тканях. Нат. Protoc. 5 , 51–66 (2010).
CAS Google Scholar
Aebi, H. Каталаза in vitro. Methods Enzymol. 105 , 121–126 (1984).
CAS Google Scholar
Ипполито, А. Дж. И Барбарик, А. К. Модифицированный метод переваривания растительной ткани азотной кислотой. Commun. Почвоведение. Завод анальный. 31 (15, 16), 2473–2482 (2000).
CAS Google Scholar
Balcke, U. G. et al. Метод UPLC-MS / MS для высокочувствительного высокопроизводительного анализа фитогормонов в тканях растений. Растительные методы 8 (47), 1–11 (2012).
Google Scholar
Livak, J. K. & Schmittgen, D. T. Анализ данных относительной экспрессии генов с использованием количественной ПЦР в реальном времени и метода 2 ΔΔCt . Методы 25 , 402–408 (2001).
CAS PubMed Central PubMed Google Scholar
Границы | Роль метаболического синдрома в раке эндометрия: обзор
Введение
Рак эндометрия — одно из самых распространенных гинекологических злокачественных новообразований.Последняя статистика рака от Американского онкологического общества показала, что в 2018 году количество новых случаев рака эндометрия в Соединенных Штатах составило 63230 человек, а количество смертей — 11350. Частота злокачественных опухолей эндометрия у женщин заняла четвертое место, а частота смерти от рака эндометрия — шестое (1). С ростом заболеваемости болезнями обмена веществ (ожирением, диабетом и гипертонией) заболеваемость раком эндометрия увеличивается и затрагивает более молодое население во всем мире.По оценкам, к 2030 году заболеваемость раком эндометрия увеличится до 42,13 на 100 000 человек в США (2). В последние годы ранняя диагностика, хирургия, лучевая терапия и химиотерапия могут значительно улучшить терапевтический эффект пациентов, но лечение ранних поражений и необходимость сохранения фертильности у поздних и рецидивирующих пациентов все еще ограничены. Клинический анализ 276 пациентов с раком эндометрия показал, что 5-летняя безрецидивная выживаемость и 5-летняя общая выживаемость составили 82.3 и 81% соответственно, а частота рецидивов и смертность от рака составили 14,5 и 15,9% соответственно (3).
Что касается патогенеза рака эндометрия, традиционная точка зрения состоит в том, что длительная непрогестероновая чрезмерная стимуляция эндометрия эстрогеном является основной причиной гиперплазии эндометрия и рака эндометрия. Эстроген может связываться с ядерным рецептором эстрогена (ER) и играть «генотипический» регулирующий эффект, регулируя транскрипцию определенных генов-мишеней.Кроме того, наши предыдущие исследования показали, что эстроген также может индуцировать приток Ca 2+ путем связывания с рецептором эстрогена, сопряженного с G-белком (GPER) на поверхности клеточной мембраны, активации кальциевого канала Cav1.3 и активации нижестоящего сигнала. путь трансдукции (MAPK / Erk) быстро, способствуя тем самым пролиферации рака эндометрия. Этот процесс не включает транскрипцию генов и синтез белка; поэтому это называется «негенетранскрипционным эффектом» (4).В настоящее время прогестерон длительного приема широко используется в клиническом лечении рака эндометрия. Однако общий эффективный уровень прогестероновой терапии первичного рака эндометрия составляет всего 50–70%, а частота рецидивов достигает 40% (5, 6). Частота объективного ответа на терапию прогестероном при распространенном и рецидивирующем раке эндометрия составляет всего 15–20% (7). Интересно, что недавние исследования показали, что уровни эстрогена в сыворотке крови у пациентов с гиперплазией эндометрия и раком эндометрия не повышены по сравнению с таковыми в нормальной контрольной группе (8).Более того, эпидемиологические исследования показали, что длительное воздействие эстрогенов у женщин в постменопаузе не увеличивает риск рака эндометрия (9). Традиционные взгляды не объясняют, почему рак эндометрия все еще встречается у женщин в постменопаузе с низким уровнем эстрогена. Эти исследования показывают, что местная чувствительность к эстрогенам, а не повышенная циркулирующая эстроген, может способствовать возникновению и развитию рака эндометрия. В то же время другие факторы, помимо эстрогена, также могут вызывать возникновение и развитие рака эндометрия.
Рак эндометрия часто ассоциируется с ожирением, диабетом и гипертонией. Эти состояния широко известны как метаболическая триада рака эндометрия. Эпидемиологические исследования показали, что риск рака эндометрия у пациентов с сахарным диабетом был в 2,12 раза выше, чем у здоровых пациентов, в то время как риск рака эндометрия у тех, кто имел избыточный вес (ИМТ ≥ 25 кг / м 2 ) был в 2,45 раза выше, чем у здоровых пациентов. в контрольной группе. Риск рака эндометрия у пациентов с ожирением и артериальной гипертензией составил 3.В 5 раз выше, чем в контрольной группе. Кроме того, рак эндометрия является одним из видов рака, наиболее тесно связанных с метаболическими заболеваниями (10). Несколько исследований показали, что метаболический синдром, вызванный ожирением, диабетом и гипертонией, тесно связан с заболеваемостью и неблагоприятным прогнозом рака эндометрия. Метаанализ шести исследований показал, что метаболический синдром тесно связан с повышенным риском рака эндометрия у женщин (относительный риск: 1,89, 95% ДИ 1,34–2.67) (11). Новое исследование показало, что метаболический синдром очень часто встречается у женщин с впервые диагностированным раком эндометрия (12). Проспективное исследование случай-контроль показало, что у женщин с впервые диагностированным раком эндометрия выше частота встречающейся гипергликемии, отношение общего холестерина к ЛПВП и три или более факторов риска сердечно-сосудистых заболеваний, чем у женщин без рака эндометрия (13). Все эти исследования показывают, что метаболический синдром тесно связан с заболеваемостью раком эндометрия.Однако точный механизм метаболического синдрома, влияющего на возникновение и развитие рака эндометрия, на сегодняшний день не определен, что может быть связано с повышением таких метаболитов, как уровень сахара в крови, инсулин, инсулиноподобный фактор роста и триглицериды в сыворотке крови (14 , 15). Динамическое взаимодействие между клетками и клеточным микроокружением играет важную роль в регулировании роста нормальных тканей и раковых клеток. Микроокружение опухолевых клеток включает опухолевые клетки и другие клетки, такие как фибробласты, лимфоциты, макрофаги, адипоциты и другие секретируемые факторы, чтобы сформировать уникальную систему микроокружения опухоли.Аномальный дисбаланс клеточного микроокружения часто приводит к онкогенезу (16). Предполагается, что молекулы, связанные с метаболическим синдромом, могут ускорять прогрессирование рака эндометрия не только за счет прямого воздействия на опухолевые клетки, но и за счет дальнейшего ремоделирования иммунного микроокружения опухолей.
В этом обзоре мы фокусируемся на метаболической микросреде рака эндометрия и суммируем ключевые молекулярные сигнальные пути ожирения, диабета и метаболического синдрома, связанного с гипертонией, влияющие на возникновение, развитие и прогноз рака эндометрия, с целью изучения новых методов ранней профилактики. , и целенаправленное лечение рака эндометрия.
Молекулярные и метаболические механизмы, лежащие в основе связи ожирения и рака эндометрия
Эпидемиологические данные показали, что ожирение тесно связано с увеличением заболеваемости различными видами рака. Некоторые ученые подтвердили причинно-следственную связь между ожирением и раком груди, построив генно-инженерную модель мышей, и показали, что ожирение тесно связано с увеличением выживаемости остаточных раковых клеток (17). Метаанализ 26 исследований, проведенных в США, показал, что каждые пять единиц увеличения индекса массы тела (ИМТ) увеличивает риск рака эндометрия на 50% [относительный риск [ОР], 1.50; 95% ДИ, 1,42–1,59] (18). В исследовании сообщается, что наличие в анамнезе бариатрической хирургии и поддержание нормального веса после операции связано со снижением риска злокачественных опухолей матки на 71 и 81% (19). Эти данные свидетельствуют о том, что ожирение может быть изменяемым фактором риска, связанным с развитием рака эндометрия. Однако механизм, с помощью которого ожирение увеличивает риск рака эндометрия, не выяснен. В настоящее время возможны следующие механизмы: пациенты с ожирением часто сопровождаются инсулинорезистентностью (гиперинсулинемия), аномальным жировым обменом (нарушения лептина, адипонектина), гипергликемией, гиперлипидемией и хроническим воспалением.Эти факторы могут способствовать возникновению и развитию опухолей (рис. 1). Чтобы изучить ключевой механизм возникновения и развития рака эндометрия, вызванного ожирением, полезно определить новую цель вмешательства для лечения пациентов с метаболическим синдромом.
Рисунок 1 . Нарушение функции жировой ткани при ожирении.
Передача сигналов эстрогена, производного адипоцитами
Эндометрий — это очень динамичная ткань, контролируемая стероидами яичников, эстрогеном и прогестероном.Длительная стимуляция эстрогенами без антагонизма прогестерона является ключевым фактором возникновения рака эндометрия. Снижение функции яичников у женщин в постменопаузе сопровождается снижением уровня гормонов. Однако женщины в постменопаузе по-прежнему подвержены раку эндометрия. Сообщалось, что полученная из жировой ткани ароматаза превращает циркулирующий андростендион в эстрадиол, что приводит к повышению уровня эстрадиола в сыворотке, который связывается с рецепторами эстрогена α и β (ERα и ERβ), что в конечном итоге приводит к привлечению факторов транскрипции, и транскрипция генов может быть активирована или репрессированные (20).Таким образом, у женщин в постменопаузе жировая ткань является основным источником биосинтеза эстрогенов. Кроме того, ожирение может привести к гиперинсулинемии, которая может снизить синтез белка, связывающего половые гормоны (SHBG), за счет увеличения биодоступности инсулиноподобного фактора роста-1 (IGF-1), что приводит к повышению уровня эстрогена. Следовательно, ожирение увеличивает риск рака эндометрия, возможно, косвенно влияя на уровень эстрогена. Метаанализ показал, что заместительная гормональная терапия (ЗГТ) может изменить ассоциацию риска рака эндометрия и ИМТ, однако менопаузальный статус и гистологический подтип не оказали существенного влияния на эти ассоциации (21).Эти данные подтверждают гипотезу о том, что гиперэстрогения является важным механизмом, лежащим в основе ассоциации ИМТ и рака эндометрия. Необходимы дополнительные исследования, чтобы изучить точный механизм, опосредующий связь между ожирением и раком эндометрия.
Резистентность к инсулину, полученному из адипоцитов
У пациентов с ожирением чрезмерное накопление жировой ткани приводит к повышенным уровням циркулирующих свободных жирных кислот и повышенной экспрессии сывороточных адипокинов, таких как лептин, висфатин и цитокины, что в конечном итоге приводит к инсулинорезистентности.Гиперинсулинемия со снижением сывороточного уровня IGF-1-связывающего белка и повышением IGF-1 чаще всего вызвана инсулинорезистентностью. Вызванное ожирением хроническое воспаление средней степени тяжести является важным фактором, приводящим к инсулинорезистентности. Воспаление, связанное с ожирением, характеризуется повышенной инфильтрацией макрофагов и повышенной экспрессией воспалительных цитокинов в жировой ткани (22). Недавно в исследованиях сообщалось, что шаперон 78 эндоплазматического ретикулума (GRP78) играет важную роль в индуцированной ожирением резистентности к инсулину, регулируя макрофаги (23).Уровень провоспалительного цитокина TGF-α в жировой ткани мышей с ожирением был значительно повышен, что было тесно связано с инсулинорезистентностью (24). Существует большое количество медиаторов воспаления, таких как C-реактивный белок (CRP), интерлейкин-6 (IL-6) и ингибитор активатора плазминогена-1 (PAI-1), которые повышены в плазме пациентов с ожирением или животных и тесно связаны с инсулинорезистентностью (25). Воспалительный ответ, вызванный ожирением, подавляет передачу сигналов инсулина в адипоцитах и гепатоцитах посредством различных сигнальных путей, включая ингибирование экспрессии субстрата 1 рецептора инсулина (IRS-1) и рецептора инсулина (IR) в сигнальных путях инсулина и ингибирование гамма-рецептора PPAR. функции, и в конечном итоге приводит к инсулинорезистентности (26, 27).Повышенный уровень инсулина и IGF-1 может стимулировать пролиферацию клеток рака эндометрия за счет связывания с рецепторами IR и IGF-1 (IGF-1R) и активации нижестоящих сигнальных путей (28).
Синергетическое взаимодействие эстрадиола и передачи сигналов инсулина
В передаче сигнала эстрогена проявляются как генотипические транскрипционные, так и нетранскрипционные эффекты: эстроген связывается с ERα в ядре, чтобы оказывать генотипические эффекты, а рецептор эстрогена GPER расположен на клеточной мембране, чтобы оказывать негенотипические транскрипционные эффекты.Исследования показали, что эстроген в сочетании с инсулином может значительно способствовать пролиферации клеток рака эндометрия по сравнению с эстрогеном или только инсулином (29). Комбинация эстрогена и диеты с высоким содержанием жиров (имитирующая инсулинорезистентность) может значительно стимулировать увеличение эндометриальных желез у мышей C57BL / 6. В этом процессе могут быть задействованы ядерный фактор, усиливающий каппа-легкую цепь активированных В-клеток (NF-κB), рецептор, активируемый пролифератором пероксисом (PPAR) и сигнальные пути LXR / RXR (30).Инсулин активирует TET1, а затем усиливает экспрессию GPER, что увеличивает чувствительность клеток рака эндометрия к эстрогену (31). Исследования показали, что эстроген и IGF-1 могут синергетически способствовать развитию опухолей у мышей, активируя сигнальные пути MAPK и AKT (32). Другие исследования показали, что эстроген может связываться с IGF-1R и оказывать негенетические транскрипционные эффекты через сигнальный путь Ras / MAPK (33). Как инсулин и IGF-1 взаимодействуют с эстрогеном в сигнальных путях, способствуя развитию рака эндометрия, требует дальнейшего изучения.
Сигнализация mTOR, полученная из жировой ткани
По сравнению с пациентами, не страдающими ожирением, активность VEGF-mTOR у больных раком эндометрия с ожирением значительно увеличилась, что позволяет предположить, что передача сигналов VEGF-mTOR, полученная из адипоцитов, является потенциальной мишенью для лечения женщин с ожирением и раком эндометрия (34). Среда, полученная из мезенхимальных стволовых клеток жировой ткани, может активировать Akt / mTOR и способствовать пролиферации и инвазии раковых клеток (35). Новый двунаправленный ингибитор PI3K и mTOR более эффективен, чем простой ингибитор mTOR (рапамицин) в подавлении рака эндометрия (36).Предполагается, что передача сигналов mTOR может быть ключевым путем, связывающим ожирение и рак эндометрия, что необходимо проверить в клинических испытаниях.
Стволовые клетки, полученные из жировой ткани
Стволовые клетки, полученные из жировой ткани, играют важную роль в микроокружении опухоли. Ожирение может способствовать трансформации стволовых клеток, полученных из жировой ткани (ASC), в связанные с раком фибробласты (CAF), тем самым способствуя пролиферации и инвазивному фенотипу раковых клеток (37). Кроме того, стволовые клетки, полученные из жировой ткани, могут способствовать метастазированию ER + клеток рака молочной железы независимо от передачи сигналов эстрогена (38).Лептин, секретируемый стволовыми клетками, полученными из жировой ткани, способствует росту и метастазированию ER + рака молочной железы за счет увеличения экспрессии рецептора ER и ароматазы (39). Сообщалось, что ASC могут сливаться с клетками рака эндометрия, а слитые клетки рака эндометрия имеют фибробластоподобный вид мезенхимального фенотипа, сопровождающийся подавлением экспрессии E-кадгерина и повышением экспрессии виментина (40). Дальнейшее изучение роли ASC в возникновении рака эндометрия поможет выяснить, как ожирение увеличивает риск рака эндометрия.
Адипокины, полученные из жиров
Адипонектин
Адипонектин — это цитокин, секретируемый в основном адипоцитами. Эпидемиологические исследования показали, что уровни адипонектина в кровообращении у пациентов с раком эндометрия снижены (41). Низкие уровни адипонектина в сыворотке тесно связаны с инсулинорезистентностью, гиперинсулинемией, ожирением и гипертонией (42). Мета-анализ показал, что уровни адипонектина в сыворотке отрицательно коррелировали с риском рака эндометрия, особенно у женщин в постменопаузе, которые не получали ЗГТ (43).В метаанализе 26 исследований для каждого увеличения адипонектина на 1 мкг / мл наблюдалось снижение суммарного относительного риска (SRR) риска рака эндометрия на 3% и снижение на 14% для каждого увеличения на 5 мкг / мл. (44). Аномальная экспрессия адипонектина в сыворотке также тесно связана с возникновением и развитием рака простаты, рака груди и рака толстой кишки (45–47). Однако противоопухолевый эффект адипонектина при раке эндометрия в основном зависит от изменения системного метаболического состояния или прямого взаимодействия с опухолевыми клетками, требующего дальнейшего изучения.
Адипонектин проявляет свое биологическое действие в основном за счет связывания с рецепторами адипонектина. Были идентифицированы три типа рецепторов адипонектина: рецептор адипонектина 1 (AdipoR1), рецептор адипонектина 2 (AdipoR2) и Т-кадгерин. AdipoR1 в основном экспрессируется в скелетных мышцах и эпителиальных клетках, а AdipoR2 наиболее широко представлен в печени (48). AMPK является важным сигнальным путем ниже адипонектина. Ось передачи сигналов адипонектин / AdipoR1 может способствовать фосфорилированию сайта AMPK Thr172, индуцируя фосфорилирование гена-супрессора опухоли LKB1.Фосфорилирование AMPK также играет важную роль в энергетическом метаболизме, активируя опухолевый супрессор TSC2 (49, 50). Исследования показали, что нет значительной разницы в экспрессии AdipoR1 и AdipoR2 в нормальных тканях эндометрия. В тканях рака эндометрия экспрессия AdipoR1 выше, чем AdipoR2. Адипонектин / AdipoRs могут ингибировать пролиферацию, адгезию и инвазивность клеток рака эндометрия, активируя нижележащую сигнальную ось LKB1-AMPK / S6 (51).Адипонектин может не только ингибировать пролиферацию и миграцию клеток рака эндометрия через сигнальный путь AMPK / mTOR / S6K1, но также может повышать чувствительность клеток рака эндометрия к инсулину через сигнальный путь AMPK / S6K1 / IRS1 (52). Адипонектин увеличивает чувствительность к инсулину в основном за счет активации активности p38MAPK. Сообщалось, что высококонсервативный сегмент из 13 остатков (ADP-1) адипонектина может способствовать транслокации транспортера глюкозы 4 (GLUT4) на клеточную мембрану, снижать уровень сахара в крови мышей db / db и способствовать секреции инсулин бета-клетками поджелудочной железы, тем самым улучшая метаболизм глюкозы и жирных кислот (53).Кроме того, антипролиферативный эффект адипонектина связан с множеством регуляторов клеточного цикла, циклином D1, D2, ERK1 / 2 и Akt.
Стоит отметить, что адипонектин не только влияет на сами опухолевые клетки, но и регулирует иммунное микроокружение опухоли. Напротив, сообщалось, что делеция адипонектина может способствовать превращению ассоциированных с опухолью макрофагов M2 в тип M1 через сигнальный путь p38MAPK, тем самым подавляя рост опухолей (54).Следовательно, с одной стороны, адипонектин может снизить частоту возникновения рака эндометрия, изменяя метаболическое состояние всего тела; с другой стороны, он может напрямую подавлять пролиферацию клеток рака эндометрия. Снижение уровня адипонектина в сыворотке крови у пациентов с ожирением тесно связано с повышенным риском рака эндометрия (44).
Висфатин
Висфатин — это инсулиноподобный адипокин, обнаруженный в последние годы. Висфатин высоко экспрессируется во многих метаболически связанных опухолях, и его повышенная экспрессия тесно связана с повышенным риском рака (55).Сообщалось, что пониженный уровень адипонектина в сыворотке или повышенный уровень висфатина являются независимыми факторами риска рака эндометрия. Соотношение висфатин: адипонектин при раке эндометрия было значительно выше, чем в контроле, что имеет определенное референтное значение для диагностики рака эндометрия (56). С увеличением ИМТ уровень висфатина у пациентов с ожирением значительно увеличивается. Повышение уровня висфатина в сыворотке тесно связано с риском инвазии миометрия (OR: 1.091; 95% ДИ: 1,021–1,166) и метастазов в лимфатические узлы (ОШ: 1,018; 95% ДИ: 1,000–1,035) рака эндометрия. Высокий уровень висфатина предполагает плохой прогноз у пациентов с раком эндометрия, который может быть потенциальной терапевтической мишенью для рака эндометрия (57, 58). Исследования показали, что висфатин может повышать экспрессию IR и субстрата инсулинового рецептора (IRS) 1/2, и он может коактивировать сигнальные пути PI3K / Akt и MAPK / ERK1 / 2 с инсулином, чтобы способствовать пролиферации и ингибировать апоптоз рака эндометрия. ячейки (59).Экзогенный висфатин может способствовать пролиферации клеток рака груди, способствуя фосфорилированию ERα и активируя эстроген-зависимые сигнальные пути (ERE) (60). Однако не сообщалось, может ли висфатин также усиливать эстроген-зависимую передачу сигналов ER и ускорять развитие рака эндометрия. Кроме того, сообщалось, что висфатин аномально экспрессируется в различных опухолях, что может увеличить риск множественных опухолей и стать потенциальным молекулярным маркером для раннего обнаружения опухолей (55).Следовательно, комбинация висфатина и адипонектина может быть маркером для ранней клинической диагностики рака эндометрия и может предоставить новые цели для клинического вмешательства.
Лептин
Лептин — важный адипокин, кодируемый геном ожирения. Лептин играет важную роль в регулировании приема пищи, потребления энергии и стимулирования роста клеток за счет объединения с рецептором лептина (ObR). Недавние исследования показали, что аномальная экспрессия лептина и передачи сигналов рецептора лептина, связанная с ожирением, играет важную роль в развитии рака груди, толстой кишки и эндометрия (61, 62).Метаанализ показал, что высокие уровни лептина могут значительно увеличить риск рака эндометрия (отношение рисков, RR = 2,55) и что высокие уровни лептина являются независимым фактором риска рака эндометрия (63). Экспрессия лептина и ObR положительно коррелирует с инвазивностью опухолей и ИМТ пациентов, но отрицательно коррелирует с гистологической степенью. Повышенная экспрессия лептина и ObR тесно связана с метастазированием в лимфатические узлы и плохим прогнозом выживания, а также с положительной экспрессией ER (64).Однако вопрос о том, может ли сигнальный путь лептина влиять на развитие рака эндометрия, влияя на классический путь передачи сигналов эстрогена, еще предстоит подтвердить. Исследования показали, что экспрессия ObR в плохо дифференцированных тканях рака эндометрия значительно выше, чем в хорошо дифференцированных тканях рака эндометрия, и лептин может ингибировать апоптоз клеток рака эндометрия, активируя сигнальный путь NIK / IKK. Повышенные уровни лептина могут влиять на полярность эпителия и способствовать злокачественной трансформации за счет сверхактивации сигнального пути PI3K / Akt (65).Лептин также способствует пролиферации и инвазии клеток рака эндометрия, активируя сигнальные пути STAT3 и ERK1 / 2, JNK. Соответственно, эта пролиферация ингибируется, когда путь JAK / STAT3 заблокирован (66). В других исследованиях сообщалось, что SNP лептина-2548 G / A может участвовать в возникновении и развитии рака эндометрия (67). Сообщалось, что лептин может активировать экспрессию STAT3-CPT1B и играет важную роль в поддержании стволовой и лекарственной устойчивости клеток рака молочной железы (68).Однако роль лептина в поддержании стволовых клеток рака эндометрия на сегодняшний день не определена и требует дальнейшего изучения. Повышенные уровни лептина предполагают наличие рака эндометрия, а уровни лептина в сыворотке могут быть эффективным инструментом для оценки клинической стадии рака эндометрия (69). Хотя повышенный уровень лептина является фактором высокого риска рака эндометрия, вопрос о том, является ли он самой важной молекулой, связанной с ожирением и раком эндометрия, требует дальнейшего исследования.
Воспалительные цитокины, полученные из жиров
Воспаление — это основная стадия онкогенеза и развития. Сообщается, что 18% случаев рака во всем мире связаны с хронической инфекцией, что подразумевает потенциальную связь между раком и воспалением. В худом состоянии баланс между адипоцитами и иммунными клетками может поддерживать нормальный обмен веществ во всем организме. Однако у людей с ожирением этот баланс выражается в ярко выраженном воспалительном микроокружении жировой ткани.Воспаление жировой ткани, связанное с ожирением, может увеличить секрецию провоспалительных факторов, вызвать системные метаболические нарушения и изменить микросреду опухолей, тем самым значительно увеличивая риск рака у людей с ожирением. Несколько исследований подтвердили, что воспалительные цитокины, связанные с ожирением, участвуют в онкогенезе.
Ил-6
IL-6 — это воспалительный цитокин, играющий важную роль во многих физиологических и патологических процессах. IL-6 тесно связан с трехкратным увеличением риска смерти у пациентов с избыточной массой тела / ожирением (70).Исследования подтвердили, что IL-6 тесно связан с возникновением множества опухолей, включая рак эндометрия. Полученный из жировой ткани IL-6 может способствовать пролиферации, инвазии и ангиогенезу клеток рака эндометрия путем активации сигнального пути JAK / STAT3 (71). Повышенный уровень IL-6 в плазме тесно связан с плохим прогнозом опухолей (72). Кроме того, эстроген может способствовать экспрессии IL-6 в раковых клетках эндометрия путем связывания с GPER на поверхности клетки (73).Сообщалось, что эстроген (E2) может способствовать экспрессии IL-6 путем связывания с ядерным рецептором ERα, а IL-6 может способствовать синтезу ароматазы путем связывания с IL-6R базальных клеток, тем самым ускоряя синтез эстрогена. и формирование контура положительной обратной связи (74). Новое исследование подтверждает, что блокирование воспалительной сигнализации, управляемой IL-6, может препятствовать распространению раковых клеток в печень (75). IL-6, вероятно, играет важную роль в развитии рака эндометрия и, по-видимому, является одним из основных механизмов, участвующих в связи между ожирением и раком.
TNF-α
TNF-α представляет собой воспалительный цитокин, секретируемый макрофагами и адипоцитами. Это важный регулятор метаболизма жировой ткани и играет важную роль в иммунной регуляции, воспалительной реакции и противоопухолевой реакции. Однако недавние исследования показали, что TNF-α также является эндогенным фактором, способствующим развитию опухоли, который может способствовать пролиферации, инвазии и метастазированию раковых клеток. Уровень TNF-α в циркулирующей крови пациентов с ожирением повышен, и повышенный уровень TNF-α тесно связан с плохим прогнозом пациентов с раком эндометрия (76).Кроме того, исследования показали, что 11 онкологических маркеров у пациентов с избыточным весом значительно выше, чем у пациентов с нормальным весом, включая ANG-2, sFASL, HB-EGF, IL-8, PLGF, TGF-α, TNF-α, uPA, VEGF-A, VEGF-C и VEGF-D (77). По сравнению с худыми мышами, мыши C57BL / 6, индуцированные диетой с высоким содержанием жиров, имели более высокие уровни сывороточных свободных жирных кислот и TNF-α и более высокое накопление макрофагов в жировой ткани (78). Исследование случай-контроль показало, что повышенные уровни TNF-α и его растворимых рецепторов (sTNFR1 и sTNFR2) были связаны с повышенным риском рака эндометрия [TNF-α-odds ratio [OR]: 1.73; sTNFR1- [OR]: 1,68; sTNFR2- [OR]: 1,53] (79). Что касается механизма TNF-α в стимулировании туморогенеза и развития, сообщалось, что хроническое воспаление, вызванное ожирением, способствует накоплению макрофагов в жировой ткани. TNF-α, высвобождаемый макрофагами M1, может способствовать метастазированию и ингибировать апоптоз клеток рака яичников путем активации сигнального пути NF-κB (80). TNF-α также является ключевым фактором, управляющим экспрессией гена ароматазы, а IL-10 может регулировать экспрессию ароматазы в жировой ткани, ингибируя сигнальный путь TNF-α (81).Более того, TNF-α может индуцировать фосфорилирование серина IRS-1 и ингибировать его запуск нижестоящих сигналов, что приводит к резистентности к инсулину. Гиперинсулинемия, вызванная инсулинорезистентностью, и IGF-1 могут дополнительно усиливать биологические эффекты TNF-α, активируя сигнальный путь TNF-α (80). TNF-α ингибировал апоптоз в раковых клетках, активируя сигнальный путь NF-κB. Хотя было идентифицировано несколько путей между TNF-α и опухолями, точный механизм связанного с ожирением TNF-α, участвующего в развитии рака эндометрия, требует дальнейшего изучения.
PAI-1
Ингибитор-1 активатора плазминогена (PAI-1) — это ингибитор протеазы, продуцируемый эндотелиальными клетками сосудов, стромальными клетками и адипоцитами в жировой ткани. Недавние исследования показали, что PAI-1 не только играет важную роль во влиянии на передачу сигналов инсулина, но также играет важную биологическую роль во влиянии на инвазию, инвазию и метастазирование опухолей, связанных с ожирением (82). PAI-1 высоко экспрессируется в тканях рака эндометрия и тесно связан с плохим прогнозом рака эндометрия (83).Исследования показали, что PAI-1 может опосредовать регуляцию транскрипции стволовых клеток, полученных из жировой ткани, при раке эндометрия (84). Следовательно, PAI-1 представляет собой потенциальную терапевтическую мишень.
Влияние ожирения на иммунную микросреду опухоли
Ожирение может способствовать секреции адипоцитами провоспалительных факторов, таких как TNF-α, IL-6 и IL-18. Эти провоспалительные цитокины могут дополнительно усиливать инфильтрацию воспалительных клеток, в основном макрофагов и Т-лимфоцитов, тем самым способствуя аномальной пролиферации и трансформации нормальных клеток (85).Исследования показали, что 16 недель аэробных тренировок и тренировок на выносливость могут снизить экспрессию воспалительных цитокинов (IL-6 и TNF-α) в жировой ткани и вызвать превращение воспалительных макрофагов M1 в противовоспалительные макрофаги M2 (86). Было обнаружено, что лептин, полученный из жировой ткани, может способствовать дифференцировке клеток Th27 и способствовать функции Т-клеток, регулируя метаболическое перепрограммирование клеток (87). Недавние исследования показали, что ожирение может увеличить инфильтрацию связанных с опухолью макрофагов, стимулировать выработку IL-1β и способствовать ангиогенезу и прогрессированию опухоли (88).Сообщалось, что макрофаги M2, инфильтрованные в микросреду рака эндометрия, могут повышать чувствительность клеток рака эндометрия к эстрогену за счет высвобождения цитокина IL17A и усиления экспрессии ERα посредством TET1-опосредованной эпигенетики (8). Эти исследования показали, что ожирение может дополнительно способствовать возникновению и прогрессированию рака эндометрия, влияя на иммунную микросреду опухолей.
Диабет 2 типа и рак эндометрия
Большинство эпидемиологических исследований предполагают, что диабет является фактором риска заболеваемости раком эндометрия; например, метаанализ 16 исследований показал, что диабет статистически значимо связан с повышенным риском рака эндометрия (сводка RR 2.10, 95% ДИ 1,75–2,53), и была более сильная связь с поправкой на возраст (ОР 2,74, 95% ДИ 1,87–4,00) (89). Кроме того, диабет тесно связан с повышенной смертностью от рака (ОР 2,09, 95% ДИ 1,31–3,35) и смертностью от причин, не связанных с раком, у женщин с раком эндометрия (90). Таким образом, эти исследования показывают, что диабет увеличивает как риск, так и смертность от рака эндометрия.
Влияние гипергликемии на рак эндометрия
Гипергликемия — важная клиническая характеристика сахарного диабета 2 типа.Системная гипергликемия создает благоприятные условия для энергетического метаболизма раковых клеток. Предыдущие исследования подтвердили, что повышенный уровень глюкозы в сыворотке может напрямую регулировать сигнальные пути, связанные с раком, особенно для удовлетворения потребностей в быстрой пролиферации раковых клеток, и может способствовать процессу перепрограммирования гликометаболизма (91). Метаболическое перепрограммирование — один из важных отличительных признаков опухолевых клеток, которые отличаются от нормальных клеток. Даже в присутствии большого количества кислорода около 80% опухолевых клеток метаболизируют глюкозу и производят АТФ в основном за счет аэробного гликолиза, также известного как эффект Варбурга.Помимо быстрого производства энергии, гликолиз также может производить большое количество промежуточных продуктов метаболизма, которые можно использовать для синтеза биологических макромолекул, необходимых для быстрого роста опухолей, включая нуклеотиды, жирные кислоты и белки.
Переносчик глюкозы (GLUT) является основным переносчиком поглощения глюкозы клетками. Когда глюкоза попадает в клетку, GLUT транспортирует аллостерическую глюкозу в клетку для поддержания высокой скорости гликолиза. Сообщалось, что высокий уровень глюкозы может способствовать экспрессии фактора роста эндотелия сосудов (VEGF) / VEGFR и процессу (эпителиально-мезенхимального перехода) EMT путем регулирования экспрессии ERα / GLUT4, тем самым способствуя пролиферации и инвазии рака эндометрия. ячейки (92).Кроме того, высокий уровень глюкозы может увеличивать активность поглощения глюкозы и гликолиза за счет регулирования путей AMPK / mTOR / S6 и MAPK, тем самым приводя к повышенной инвазивности клеток рака эндометрия. Более того, высокий уровень глюкозы может способствовать пролиферации клеток рака эндометрия за счет активации экспрессии STAT3, которая может подавляться метформином (93). Активность метаболизма глюкозы тесно связана с концентрацией глюкозы вне клетки. Однако неясно, как клетки воспринимают внешние уровни глюкозы и регулируют активность пути гликолиза.AMPK, аденилат-активируемая протеинкиназа, является ключевым белком для восприятия концентрации внеклеточной глюкозы (94). Сообщалось, что высокие уровни внеклеточной глюкозы регулируют уровень белка CARM1 за счет снижения фосфорилирования AMPK, тем самым ингибируя метилирование GAPDH, что дополнительно способствует активности фермента GAPDH и пути гликолиза (95). Это исследование раскрывает механизм, с помощью которого клетки воспринимают уровни глюкозы во внеклеточной среде и регулируют скорость гликолиза, помогая выяснить механизм, с помощью которого раковые клетки воспринимают и используют глюкозу.
Изоферменты пируваткиназы M2 (PKM2) — ключевой метаболический фермент, который способствует гликолизу и играет важную роль в онкогенезе благодаря эффекту Варбурга. Сообщалось, что высокий уровень глюкозы может способствовать аномальной экспрессии PKM2. Сверхэкспрессия PKM2 может способствовать накоплению промежуточных продуктов гликолиза (пирувата и молочной кислоты), обеспечивать предшественников для синтеза биомакромолекул и приводить к пролиферации клеток и онкогенезу (96). Молочная кислота является ключевым метаболитом гликолиза в раковых клетках.Накопление внеклеточной молочной кислоты оказывает важное влияние на метаболизм раковых клеток и трансформацию нераковых клеток в раковые, включая метаболическое перепрограммирование, опухолевое воспаление и ангиогенез. Недавние исследования показали, что молочная кислота, продуцируемая гликолизом раковых клеток, может подавлять функцию макрофагов через механизм, опосредованный гипоксией, индуцировать трансформацию противоопухолевых макрофагов типа M1 в M2 и способствовать инвазии и миграции рака. ячейки (97).Интересно, что макрофаги M2 являются преобладающими опухоль-ассоциированными макрофагами при раке эндометрия и играют важную роль в возникновении и развитии рака эндометрия (98). Транспортер монокарбоксилата 1 (MCT1) является важным белком для поглощения молочной кислоты и пирувата клетками, в то время как MCT4 является важным белком для клеточного транспорта молочной кислоты и пирувата, которые играют важную роль в регуляции метаболизма молочной кислоты. Сообщалось, что MCT1 был независимым прогностическим биомаркером рака эндометрия (99).Недавнее исследование показало, что использование ингибиторов переносчиков монокарбоновых кислот (МСТ) может обратить вспять ингибирование молочной кислоты на лизосомах макрофагов (100). Следовательно, ингибирование MCT1 может иметь потенциал для лечения рака эндометрия. Предполагается, что усиление активности гликолиза и метаболитов, таких как молочная кислота, при раке эндометрия в среде с высоким содержанием глюкозы может привести к подкислению микроокружения опухоли. Кислая среда может восприниматься связанными с опухолью макрофагами, которые зависят от пути транспортера монокарбоновой кислоты 1 (MCT1), чтобы вызвать трансформацию макрофагов из M1 в M2, тем самым ускоряя прогрессирование рака эндометрия.Эти исследования показали, что лечение гипергликемии или нацеливание на гликометаболизм может быть потенциальной терапевтической стратегией при раке эндометрия.
Влияние резистентности к инсулину на рак эндометрия
Инсулинорезистентность и гиперинсулинемия — важные характеристики ожирения и диабета. Экспрессия инсулина и IGF-1 была значительно увеличена у пациентов с диабетом, а высокие уровни инсулина были независимым фактором рака эндометрия (28, 101, 102). Высокий уровень инсулина и IGF-1/2 у пациентов с диабетом может ускорить превращение андростендиона в эстроген под действием ароматазы и повысить уровень эстрогена за счет ингибирования синтеза SHBG.Длительная стимуляция эстрогенами без антагонизма прогестерона может вызвать дисплазию эндометрия или даже злокачественную трансформацию (103). Сообщалось, что после поправки на ИМТ, возраст и гистологический тип высокая экспрессия IR / IGF-1R тесно связана с прогностическими факторами высокого риска, такими как прогрессирование рака эндометрия и инфильтрация лимфатических узлов (104).
Инсулин может активировать фосфоинозитид-3-киназу (PI3K) / AKT или митоген-активированную протеинкиназу (MAPK) / сигнальный путь регулируемой киназы ERK с внеклеточными сигналами посредством связывания с IR / IGF-1R для стимулирования EMT рака эндометрия, что приводит к увеличению пролиферация и инвазия клеток рака эндометрия, подавляет апоптоз раковых клеток и способствует ангиогенезу опухолей (105).Кроме того, инсулин может усиливать экспрессию фактора роста эндотелия сосудов, тем самым стимулируя ангиогенез, который тесно связан с возникновением и развитием опухолей (106). Недавние исследования показали, что инсулино-зависимое фосфорилирование инициирует метаболизм глюкозы до транспорта глюкозы; таким образом, метаболизм глюкозы направлен в определенное направление гликолиза (107). Таким образом, передача сигналов инсулина играет ключевую роль в перепрограммировании гликометаболизма, а инсулинорезистентность может способствовать возникновению и развитию рака эндометрия посредством непрямого пути или прямого пути лиганд-рецептор.
IR-A имеет высокое сродство к инсулину и IGF-II, но связывает IGF-I с низким сродством. IGF-1R имеет высокое сродство к IGF. Однако блокирование IGF-IR и IR не полностью предотвращает стимуляцию роста инсулиноподобного фактора роста или инсулина на раковых клетках, предполагая, что другие рецепторы могут участвовать в сложных системах передачи сигналов. Наши предыдущие исследования обнаружили и впервые подтвердили экспрессию Hybrid-R при раке эндометрия, который может способствовать пролиферации и подавлять апоптоз клеток рака эндометрия через сигнальный путь MAPK / ERK (22).Ожидается, что Hybrid-R предоставит новую терапевтическую цель и стратегию точного лечения пациентов с раком эндометрия с инсулинорезистентностью и гиперинсулинемией.
Метформин в лечении рака эндометрия
Метформин, сенсибилизатор инсулина, считается потенциальным противоопухолевым препаратом. Наши предыдущие клинические исследования показали, что метформин в сочетании с лечением прогестероном может значительно улучшить эффективность у пациентов с раком эндометрия с плохим лечением прогестероном (108).Сообщалось, что гипергликемия плазмы и высокие уровни IGF-1 у пациентов с раком эндометрия могут быть купированы обычными дозами метформина (109). Метформин может значительно ингибировать пролиферацию клеток рака эндометрия, что может быть связано с активацией передачи сигналов AMPK и ингибированием пути передачи сигналов mTOR (110). Напротив, некоторые исследования показали, что метформин не снижает риск рака эндометрия и не может улучшить общую выживаемость пациентов (111).Соответственно, исследования показали, что метформин не влияет на PI3K-Akt-mTOR и сигнальные пути инсулина и не влияет на потерю веса (112). Большое клиническое значение имеет дальнейшее изучение механизма аномального метаболизма глюкозы, способствующего развитию рака эндометрия, выявление ключевой молекулы рака эндометрия, вызываемой метаболическими заболеваниями, и поиск новых эффективных профилактических или противораковых препаратов, которые могут заменить метформин.
Другие типы заболеваний метаболического синдрома и рака эндометрия
Дислипидемия тесно связана с заболеваемостью различными видами рака (113).Сообщалось, что у пациентов с гипергликемией, гиперлипидемией и гипертонией вероятность развития рака эндометрия в два раза выше, чем у нормальных людей (114). ИМТ пациентов положительно коррелировал с уровнями пальмитиновой кислоты, олеиновой кислоты и стеариновой кислоты в сыворотке крови. Увеличение свободных жирных кислот у пациентов с ожирением может косвенно способствовать пролиферации клеток рака эндометрия за счет повышения уровня эстрадиола (115). ИМТ пациентов с раком эндометрия был значительно выше, чем у здоровых людей.В настоящее время существует множество исследований взаимосвязи между раком эндометрия, ожирением и диабетом, но мало исследований, посвященных взаимосвязи между раком эндометрия и гипертонией и нарушением липидов. Возможные механизмы аномальных липидов крови, связанные с риском рака эндометрия, следующие: активация путей жирных кислот и аминогексозы приводит к производству активных форм кислорода (АФК) в митохондриях, что вызывает окислительный стресс в клетках. Чрезмерная агрегация кластеров АФК взаимодействует с липидами, белками и ДНК в клетках, вызывая изменения мембранных и ферментативных функций, вызывая повреждение клеток и, в конечном итоге, приводя к онкогенезу (116).Недавние исследования показали, что уровень холестерина в сыворотке повышен у людей с ожирением. Холестерин активирует транскрипционную активность клеток рака эндометрия через ER-зависимый путь и способствует пролиферации клеток рака эндометрия (117). Более того, недавние исследования показали, что повышенный уровень циркулирующих свободных жирных кислот может быть важным фактором в связывании ожирения и туморогенеза, что может способствовать пролиферации и инвазии клеток рака молочной железы через передачу сигналов ERα и сигнальный путь mTOR (118).Сообщалось, что A-FABP, высвобождаемый из жировой ткани, может способствовать сухости и инвазивности клеток рака груди (119). Взятые вместе, аномальный липидный обмен, особенно повышенный уровень свободных жирных кислот, тесно связан с прогрессированием рака эндометрия. Могут ли жирные кислоты участвовать в возникновении и развитии рака эндометрия посредством ER-зависимого пути, еще предстоит тщательно охарактеризовать.
Гипотиреоз — это тип синдрома пониженного метаболизма, вызванный снижением синтеза и секреции гормонов щитовидной железы или неадекватными физиологическими эффектами.Бринтон и др. указали, что ЭК связана с предыдущими диагнозами заболеваний щитовидной железы (ОР = 1,52, 95% ДИ 1,17–1,98) (120). Сообщалось, что частота гипотиреоза у пациентов с ЭК значительно увеличивается. Уровень ТТГ в сыворотке до лечения является независимым фактором риска плохого прогноза ЭК (121). Кроме того, сообщалось, что повышенный уровень ТТГ увеличивает частоту рассеянного склероза (122). В настоящее время исследований функции щитовидной железы и рака эндометрия мало. Несколько исследований показали, что гипотиреоз тесно связан с рассеянным склерозом, СПКЯ, повышенным уровнем лептина в сыворотке и дислипидемией.Гипотиреоз может способствовать возникновению и развитию рака эндометрия, косвенно увеличивая факторы риска рака эндометрия или за счет прямого взаимодействия (123). Исследования, изучающие взаимосвязь между функцией щитовидной железы (уровнем ТТГ) и раком эндометрия, дадут новое понимание механизма рака эндометрия.
Заключение
Метаболический синдром — сложное заболевание, определяемое кластером метаболических факторов риска, который включает инсулинорезистентность, гиперинсулинемию, нарушение толерантности к глюкозе, сахарный диабет 2 типа, дислипидемию и висцеральное ожирение.Ожирение, диабет и гипертония — это метаболическая триада рака эндометрия. У женщин с впервые диагностированным раком эндометрия был очень высокий уровень распространенности метаболического синдрома. В этой статье мы рассматриваем возможные пути, напрямую связывающие метаболический синдром с раком (рис. 2). Связанная с ожирением инсулинорезистентность, уровни лептина и лактона тесно связаны с возникновением и развитием рака эндометрия. Снижение уровня адипонектина в сыворотке и усиление хронического воспаления у пациентов с ожирением являются важными факторами повышения риска рака эндометрия.Ожирение и диабет имеют много общих патологических характеристик: инсулинорезистентность (гиперинсулинемия), аномальный жировой обмен (повышенный лептин, пониженный адипонектин), гипергликемия, гиперлипидемия и хроническое воспаление. Во многих исследованиях сообщается, что эти характеристики могут способствовать возникновению и развитию рака эндометрия, напрямую воздействуя на опухолевые клетки или регулируя микросреду опухоли. Следовательно, существует острая необходимость в вмешательстве при хронических заболеваниях, связанных с метаболическим синдромом, для снижения заболеваемости раком эндометрия.Комментарий MacKintosh et al. указанное ожирение, перенесшее бариатрическую операцию или медикаментозное снижение веса, может снизить риск рака эндометрия, а гиперпластические аномалии эндометрия могут быть обратимы за счет потери веса (124). В исследованиях оценивали влияние хирургического вмешательства при ожирении и похудании на морфологию эндометрия и молекулярные сигнальные пути при раке эндометрия. Было обнаружено, что циркулирующие биомаркеры инсулинорезистентности (HbA1c, HOMA-IR) и воспаления (hsCRP, IL-6) снижались, а биомаркеры репродуктивной системы (ЛГ, ФСГ, ГСПГ) значительно увеличивались (125).C-пептид, инсулин, C-реактивный белок, лептин, IL-1Ralpha и IL-6 значительно снизились, в то время как SHBG, IGFBP1 и адипонектин значительно увеличились при вмешательствах по снижению веса при раке эндометрия (126). Таким образом, бариатрическая хирургия может снизить риск рака эндометрия за счет уменьшения воспаления, вызванного ожирением. Тем не менее, роль бариатрической хирургии в лечении гиперплазии эндометрия все еще требует неопровержимых или убедительных доказательств. Недавно было предложено семь маркеров на основе чувствительного к ИМТ пути инсулинорезистентности, adipoR1, adipoR2, ObR, IRβ, IRS-1, IGF-1R и IGF-2R, для разработки новой системы молекулярного типирования для рака эндометрия.Однако эффективных систем молекулярного типирования и маркеров молекулярного типирования для рака эндометрия не обнаружено (127). В исследовании была предложена прагматическая модель прогнозирования риска рака эндометрия, которая включала ожирение, репродуктивную функцию, инсулинорезистентность и генетический риск. Эта модель играет важную роль в выявлении лиц с высоким риском рака эндометрия и в руководстве профилактическим лечением конкретных целей заболевания (128). За исключением ожирения, репродуктивной функции, инсулинорезистентности и генетического риска, следует также учитывать другие факторы высокого риска рака эндометрия, связанные с метаболическим синдромом, упомянутые в тексте, и требуется большая проспективная когорта бессимптомных женщин.Точно так же исследования, включающие биомаркеры (адипонектин, эстрадиол, антагонист рецептора интерлейкина-1, фактор некроза опухоли и триглицерид) в модель прогнозирования риска рака эндометрия, показали, что они могут незначительно улучшить прогностическую способность рака эндометрия (129). Несмотря на различные исследования, сообщалось о новых молекулярных маркерах, но на сегодняшний день надежные молекулярные маркеры не применялись для клинического молекулярного типирования. Наше недавнее исследование показало, что общий кальций сыворотки может быть более чувствительным параметром метаболического синдрома, чем гиперлипидемия у пациентов с эндометриоидным раком (130).
Рисунок 2 . Возможные пути прямой связи метаболического синдрома с раком эндометрия.
Следовательно, необходимо провести скрининг новых маркеров, основанных на системных метаболомных изменениях, и выявить новые методы молекулярного типирования и прогностические модели риска рака эндометрия, основанные на метаболических изменениях в сыворотке крови. При частом применении протеомики, метаболомики и транскриптомов роль ключевых молекул в идентификации заболеваний, связанных с метаболическим синдромом, при раке эндометрия имеет большое значение для ранней профилактики и лечения рака эндометрия.В заключение, рак эндометрия — это тип опухоли, связанной с метаболическим заболеванием. Ожидается, что выяснение конкретных ролей и механизмов заболеваний, связанных с метаболическим синдромом, при раке эндометрия станет новой целью для ранней профилактики и лечения рака эндометрия. Хотя изучение связи между метаболическим синдромом и раком может обеспечить эффективную терапевтическую мишень для рака эндометрия, улучшение образа жизни по-прежнему является наиболее важным компонентом в предотвращении заболеваемости и смертности от рака эндометрия, связанного с метаболическим синдромом.Дальнейшие in vivo и клинические исследования необходимы для изучения терапевтического воздействия на метаболическое микроокружение при раке эндометрия, связанном с метаболическим синдромом.
Авторские взносы
XY и JW написали и утвердили окончательную версию этой рукописи.
Финансирование
Эта работа была поддержана Национальным фондом естественных наук Китая (гранты № 81672571 и 81874108), Специальными проектами по усилению фундаментальных исследований Пекинского университета (грант №BMU2018JC005) и Национальная программа исследований и разработок ключевых технологий (грант № 2015BAI13B06).
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Список литературы
2. Шейх М.А., Альтхаус А.Д., Фриз К.Э., Суассон С., Эдвардс Р.П., Велберн С. и др. Прогнозы рака эндометрия в США до 2030 года: стоит ли нам беспокоиться? Future Oncol. (2014) 10: 2561–8. DOI: 10.2217 / fon.14.192
PubMed Аннотация | CrossRef Полный текст | Google Scholar
3. Техерисо-Гарсия А., Хименес-Лопес Дж. С., Муньос-Гонсалес Дж. Л., Бартоломе-Сотильос С., Маркета-Маркес Л., Лопес-Гонсалес Г. и др. Общая выживаемость и выживаемость без заболеваний при раке эндометрия: прогностические факторы у 276 пациентов. Onco Targets Ther. (2013) 9: 1305–13. DOI: 10.2147 / OTT.S51532
PubMed Аннотация | CrossRef Полный текст | Google Scholar
4.Хао Дж, Бао Х, Цзинь Б., Ван Х, Мао З, Ли Х и др. Субъединица альфа-1D канала Ca2 + способствует пролиферации и миграции клеток рака эндометрия, опосредованной 17-бета-эстрадиолом, через рецептор эстрогена, связанный с G-белком. FASEB J. (2015) 29: 2883–93. DOI: 10.1096 / fj.14-265603
PubMed Аннотация | CrossRef Полный текст | Google Scholar
7. Thigpen JT, Brady MF, Alvarez RD, Adelson MD, Homesley HD, Manetta A, et al. Пероральный прием медроксипрогестерона ацетата при лечении запущенной или рецидивирующей карциномы эндометрия: исследование зависимости зависимости от дозы, проведенное Группой гинекологической онкологии. J Clin Oncol. (1999) 17: 1736–44. DOI: 10.1200 / JCO.1999.17.6.1736
PubMed Аннотация | CrossRef Полный текст | Google Scholar
8. Нин Ц., Се Б., Чжан Л., Ли Ц., Шань В., Ян Б. и др. Проникающие макрофаги индуцируют экспрессию ERalpha посредством опосредованного IL17A эпигенетического механизма, повышая чувствительность клеток рака эндометрия к эстрогену. Cancer Res. (2016) 76: 1354–66. DOI: 10.1158 / 0008-5472.CAN-15-1260
PubMed Аннотация | CrossRef Полный текст | Google Scholar
9.Cherry N, McNamee R, Heagerty A, Kitchener H, Hannaford P. Долгосрочная безопасность не имеющего сопротивления эстрогена, используемого женщинами, перенесшими инфаркт миокарда: 14-летнее наблюдение рандомизированного контролируемого исследования ESPRIT. BJOG. (2014) 121: 700–5. Обсуждение 5. doi: 10.1111 / 1471-0528.12598
PubMed Аннотация | CrossRef Полный текст | Google Scholar
11. Эспозито К., Чиодини П., Капуано А., Белластелла Г., Майорино М. И., Джульяно Д. Метаболический синдром и рак эндометрия: метаанализ. Эндокринная . (2014) 45: 28–36. DOI: 10.1007 / s12020-013-9973-3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
12. Китсон С.Дж., Линдси Дж., Сивалингам В.Н., Раттер М.К., Кросби Э.Дж. Высокая распространенность метаболического синдрома у женщин с впервые диагностированным раком эндометрия. Gynecol Oncol Rep. (2018) 26: 109–10. DOI: 10.1016 / j.gore.2018.08.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
13. Китсон С.Дж., Линдси Дж., Сивалингам В.Н., Лант М., Райан Н.Дж., Эдмондсон Р.Дж. и др.Неизвестное бремя факторов риска сердечно-сосудистых заболеваний у женщин с впервые диагностированным раком эндометрия: проспективное исследование случай-контроль. Gynecol Oncol. (2018) 148: 154–60. DOI: 10.1016 / j.ygyno.2017.11.019
PubMed Аннотация | CrossRef Полный текст | Google Scholar
14. Trabert B, Wentzensen N, Felix AS, Yang HP, Sherman ME, Brinton LA. Метаболический синдром и риск рака эндометрия в Соединенных Штатах: исследование в связанной базе данных SEER-medicare. Биомаркеры эпидемиологии рака Пред. (2015) 24: 261–7. DOI: 10.1158 / 1055-9965.EPI-14-0923
PubMed Аннотация | CrossRef Полный текст | Google Scholar
15. Nead KT, Sharp SJ, Thompson DJ, Painter JN, Savage DB, Semple RK, et al. Доказательства причинной связи между инсулинемией и раком эндометрия: менделевский рандомизационный анализ. J Natl Cancer Inst. (2015) 107: djv178. DOI: 10.1093 / jnci / djv178
PubMed Аннотация | CrossRef Полный текст | Google Scholar
16.Куни Т., де Гердер В., Барлье А., Хофланд Л.Дж. Роль микросреды опухоли в нейроэндокринных опухолях пищеварительной системы. Endocr Relat Cancer. (2018) 25: R519–44. DOI: 10.1530 / ERC-18-0025
PubMed Аннотация | CrossRef Полный текст | Google Scholar
17. Эккер Б.Л., Ли Дж.Й., Стернер С.Дж., Соломон А.С., Пант Д.К., Шен Ф. и др. Влияние ожирения на рецидивы рака груди и минимальную остаточную болезнь. Breast Cancer Res. (2019) 21:41. DOI: 10.1186 / s13058-018-1087-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
18.Всемирный фонд исследований рака / Американский институт исследований рака. Постоянное обновление отчета по проекту. Питание, питание, физическая активность и профилактика рака эндометрия . (2013). Доступно в Интернете по адресу: http://www.dietandcancerreport.org (по состоянию на 1 августа 2019 г.).
Google Scholar
19. Ward KK, Roncancio AM, Shah NR, Davis MA, Saenz CC, McHale MT, et al. Бариатрическая хирургия снижает риск злокачественного новообразования матки. Gynecol Oncol. (2014) 133: 63–6.DOI: 10.1016 / j.ygyno.2013.11.012
PubMed Аннотация | CrossRef Полный текст | Google Scholar
21. Кросби Э.Дж., Звален М., Китченер Х.С., Эггер М., Ренехан АГ. Индекс массы тела, заместительная гормональная терапия и риск рака эндометрия: метаанализ. Биомаркеры эпидемиологии рака Пред. (2010) 19: 3119–30. DOI: 10.1158 / 1055-9965.EPI-10-0832
PubMed Аннотация | CrossRef Полный текст | Google Scholar
22. Йе Дж, Макгиннесс ОП. Воспаление при ожирении — это не все плохо: данные исследований на животных и людях. Am J Physiol Endocrinol Metab. (2013) 304: E466–77. DOI: 10.1152 / ajpendo.00266.2012
CrossRef Полный текст | Google Scholar
23. Ким Дж. Х., Ли Е., Фридлин Р. Х., Сук С., Юнг Д. Ю., Дагдевирен С. и др. Шаперон эндоплазматического ретикулума GRP78 регулирует функцию макрофагов и инсулинорезистентность при ожирении, вызванном диетой. FASEB J. (2018) 32: 2292–304. DOI: 10.1096 / fj.201701017R
PubMed Аннотация | CrossRef Полный текст | Google Scholar
24.Сзето Х.Х., Лю С., Сунг Й., Алам Н., Пруски Г.Т., Сешан С.В. Защита митохондрий предотвращает гломерулопатию, вызванную высоким содержанием жиров, и повреждение проксимальных канальцев. Kidney Int. (2016) 90: 997–1011. DOI: 10.1016 / j.kint.2016.06.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
26. Перротта Ф., Нигро Э., Моллика М., Костильола А., Д’Аньяно В., Даниэле А. и др. Легочная гипертензия и ожирение: фокус на адипонектин. Int J Mol Sci. (2019) 20: E912.DOI: 10.3390 / ijms20040912
PubMed Аннотация | CrossRef Полный текст | Google Scholar
27. Лю З., Ван Н., Ма И, Вэнь Д. Гидрокситирозол улучшает ожирение и резистентность к инсулину, модулируя микробиоту кишечника у мышей с ожирением, вызванным диетой с высоким содержанием жиров. Front Microbiol. (2019) 10: 390. DOI: 10.3389 / fmicb.2019.00390
PubMed Аннотация | CrossRef Полный текст | Google Scholar
29. Тянь В., Тэн Ф., Чжао Дж., Гао Дж., Гао С., Сунь Д. и др. Эстроген и инсулин синергетически способствуют прогрессированию рака эндометрия 1 типа. Cancer Biol Ther. (2017) 18: 1000–10. DOI: 10.1080 / 15384047.2017.1394547
PubMed Аннотация | CrossRef Полный текст | Google Scholar
30. Cheng Y, Lv Q, Xie B, Yang B, Shan W., Ning C, et al. Эстроген и диета с высоким содержанием жиров вызывали изменения в профиле транскриптома эндометрия мышей C57BL / 6. Endocr Connect. (2018) 7: 36–46. DOI: 10.1530 / EC-17-0315
PubMed Аннотация | CrossRef Полный текст | Google Scholar
31. Xie BY, Lv QY, Ning CC, Yang BY, Shan WW, Cheng YL и др.Путь TET1-GPER-PI3K / AKT участвует в инсулино-управляемой пролиферации клеток рака эндометрия. Biochem Biophys Res Commun. (2017) 482: 857–62. DOI: 10.1016 / j.bbrc.2016.11.124
PubMed Аннотация | CrossRef Полный текст | Google Scholar
32. Тан Х, Ляо Й, Сюй Л., Чжан Ц., Лю З., Дэн И и др. Эстроген и инсулиноподобный фактор роста 1 синергетически способствуют развитию аденокарциномы легких у мышей. Int J Cancer. (2013) 133: 2473–82. DOI: 10.1002 / ijc.28262
PubMed Аннотация | CrossRef Полный текст | Google Scholar
33. Каманга-Солло Е., Белый ME, Хэтэуэй М.Р., Чанг К.Ю., Джонсон Б.Дж., Дейтон В.Р. Роль IGF-I и рецепторов эстрогена, андрогена и IGF-I в пролиферации культивируемых сателлитных клеток крупного рогатого скота, стимулированной эстрадиол-17бета и тренболона ацетатом. Domest Anim Endocrinol. (2008) 35: 88–97. DOI: 10.1016 / j.domaniend.2008.02.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
34.Саху С.С., Ломбард Д.М., Иус Й., О’Салливан Р., Вуд Л.Г., Нахар П. и др. Передача сигналов VEGF-mTOR, полученная из жировой ткани, способствует гиперплазии эндометрия и раку: последствия для женщин с ожирением. Mol Cancer Res. (2018) 16: 309–21. DOI: 10.1158 / 1541-7786.MCR-17-0466
PubMed Аннотация | CrossRef Полный текст | Google Scholar
35. Ко Дж. Х., Ум Дж. Й., Ли С. Г., Ян В. М., Сетхи Дж., Ан К. С.. Кондиционированные среды из адипоцитов способствуют пролиферации, миграции и инвазии в клетки меланомы и колоректального рака. J. Cell Physiol. (2019) 234: 18249–61. DOI: 10.1002 / jcp.28456
PubMed Аннотация | CrossRef Полный текст | Google Scholar
36. Чен Дж, Чжао К.Н., Ли Р., Шао Р., Чен С. Активация пути PI3K / Akt / mTOR и двойных ингибиторов PI3K и mTOR при раке эндометрия. Curr Med Chem. (2014) 21: 3070–80. DOI: 10.2174 / 0
7321666140414095605PubMed Аннотация | CrossRef Полный текст | Google Scholar
37. Стронг А.Л., Пей Д.Т., Херст К.Г., Гимбл Дж.М., Буроу М.Э., Баннелл Б.А.Ожирение усиливает преобразование стромальных / стволовых клеток, полученных из жировой ткани, в фибробласты, связанные с карциномой, что приводит к пролиферации раковых клеток и прогрессированию до инвазивного фенотипа. Stem Cells Int. (2017) 2017:
02. DOI: 10.1155 / 2017/
02
PubMed Аннотация | CrossRef Полный текст | Google Scholar
38. Сабол Р.А., Бейли А., Джакомелли П., Уайз Р.М., Харрисон МАА, О’Доннелл Б.А. и др. Стволовые жировые клетки, измененные ожирением, способствуют метастазированию рака молочной железы ER (+) через эстроген-независимые пути. Int J Mol Sci. (2019) 20: E1419. DOI: 10.3390 / ijms20061419
PubMed Аннотация | CrossRef Полный текст | Google Scholar
39. Strong AL, Strong TA, Rhodes LV, Semon JA, Zhang X, Shi Z, et al. Связанные с ожирением изменения в биологии жировых стволовых клеток опосредуют усиленный туморогенез эстроген-зависимыми путями. Breast Cancer Res. (2013) 15: R102. DOI: 10.1186 / bcr3569
PubMed Аннотация | CrossRef Полный текст | Google Scholar
40.Ли М., Ли Х, Чжао Л., Чжоу Дж., Ченг И, Сюй Б. и др. Спонтанное образование онкогенных гибридов между стромальными клетками человеческого сальника, полученными из жировой ткани, и клетками рака эндометрия увеличивало подвижность и гетерогенность раковых клеток. Cell Cycle. (2019) 18: 320–32. DOI: 10.1080 / 15384101.2019.1568743
PubMed Аннотация | CrossRef Полный текст | Google Scholar
41. Rzepka-Gorska I, Bedner R, Cymbaluk-Ploska A, Chudecka-Glaz A. Сывороточный адипонектин в отношении рака эндометрия и гиперплазии эндометрия с атипией у женщин с ожирением. Eur J Gynaecol Oncol. (2008) 29: 594–7.
PubMed Аннотация | Google Scholar
42. Аггелусси С., Теодору А.А., Пашалис В., Николаидис М.Г., Фатурос И.Г., Оволаби Е.О. и др. Уровни адипоцитокинов у детей: эффекты ожирения и тренировок. Pediatr Exerc Sci. (2012) 24: 461–71. DOI: 10.1123 / pes.24.3.461
PubMed Аннотация | CrossRef Полный текст | Google Scholar
43. Zeng F, Shi J, Long Y, Tian H, Li X, Zhao AZ, et al. Адипонектин и рак эндометрия: систематический обзор и метаанализ. Cell Physiol Biochem. (2015) 36: 1670–8. DOI: 10.1159 / 000430327
PubMed Аннотация | CrossRef Полный текст | Google Scholar
45. Карнати Х.К., Паниграхи М.К., Ли Й., Твиди Д., Грейг Н.Х. Адипонектин как потенциальная терапевтическая мишень при раке простаты. Curr Pharm Des. (2017) 23: 4170–9. DOI: 10.2174/1381612823666170208123553
PubMed Аннотация | CrossRef Полный текст | Google Scholar
47. Нигро Э., Скеттино П., Полито Р., Скудьеро О, Монако М.Л., Де Пальма Г.Д. и др.Адипонектин и рак толстой кишки: доказательства ингибирующего воздействия на жизнеспособность и миграцию линий колоректальных клеток человека. Mol Cell Biochem. (2018) 448: 125–35. DOI: 10.1007 / s11010-018-3319-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
48. Панно М.Л., Наимо Г.Д., Спина Э., Андо С., Мауро Л. Различные молекулярные сигналы, поддерживающие действие адипонектина при раке груди. Curr Opin Pharmacol. (2016) 31: 1–7. DOI: 10.1016 / j.coph.2016.08.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
50.Ван И, Сюй В., Янь З., Чжао В., Ми Дж, Ли Дж и др. Метформин вызывает аутофагию и остановку клеточного цикла в фазе G0 / G1 при миеломе, воздействуя на пути AMPK / mTORC1 и mTORC2. J Exp Clin Cancer Res. (2018) 37:63. DOI: 10.1186 / s13046-018-0731-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
51. Мун Х.С., Чемберленд Дж. П., Аронис К., Целени-Балафута С., Манцорос К. С.. Прямая роль адипонектина и рецепторов адипонектина при раке эндометрия: исследования in vitro, и ex vivo, на людях. Mol Cancer Ther. (2011) 10: 2234–43. DOI: 10.1158 / 1535-7163.MCT-11-0545
PubMed Аннотация | CrossRef Полный текст | Google Scholar
52. Цай З.Ф., Дэн Л., Ван М.М., Чжан Дж.К., Ли Л. [Эффект путей AMPK / mTOR / S6K1 и инсулино-сенсибилизирующий эффект адипонектина в раковых клетках эндометрия]. Чжунхуа Фу Чан Кэ За Чжи. (2018) 53: 554–60. DOI: 10.3760 / cma.j.issn.0529-567x.2018.08.008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
53.Сайед М., Гаутам С., Верма Д.П., Афшан Т., Кумари Т., Шривастава А.К. и др. Полученный из коллагенового домена короткий пептид адипонектина активирует сигнальные пути APPL1 и AMPK и улучшает метаболизм глюкозы и жирных кислот. J. Biol Chem. (2018) 293: 13509–23. DOI: 10.1074 / jbc.RA118.001801
PubMed Аннотация | CrossRef Полный текст | Google Scholar
54. Peng J, Tsang JY, Ho DH, Zhang R, Xiao H, Li D, et al. Модулирующие эффекты адипонектина на поляризацию опухоль-ассоциированных макрофагов. Int J Cancer. (2015) 137: 848–58. DOI: 10.1002 / ijc.29485
PubMed Аннотация | CrossRef Полный текст | Google Scholar
55. Mohammadi M, Mianabadi F, Mehrad-Majd H. Уровни циркулирующего висфатина и риск рака: систематический обзор и метаанализ. J. Cell Physiol. (2019) 234: 5011–22. DOI: 10.1002 / jcp.27302
PubMed Аннотация | CrossRef Полный текст | Google Scholar
56. Wang Z, Gao S, Sun C, Li J, Gao W, Yu L. Клиническое значение уровней адипонектина и висфатина в сыворотке при раке эндометрия. Int J Gynaecol Obstet . (2019) 145: 34–9. DOI: 10.1002 / ijgo.12772
PubMed Аннотация | CrossRef Полный текст | Google Scholar
57. Ильхан Т.Т., Кебапджилар А., Йилмаз С.А., Ильхан Т., Керимоглу О.С., Пекин А.Т. и др. Связь уровней висфатина и резистина в сыворотке с раком эндометрия и факторами, связанными с его прогнозом. Азиатский Pac J Cancer Prev. (2015) 16: 4503–8. DOI: 10.7314 / APJCP.2015.16.11.4503
PubMed Аннотация | CrossRef Полный текст | Google Scholar
58.Cymbaluk-Ploska A, Chudecka-Glaz A, Pius-Sadowska E, Sompolska-Rzechula A, Machalinski B, Menkiszak J. Циркулирующий уровень висфатина в сыворотке крови у пациентов с раком эндометрия. Biomed Res Int. (2018) 2018: 8576179. DOI: 10.1155 / 2018/8576179
PubMed Аннотация | CrossRef Полный текст | Google Scholar
59. Ван И, Гао Ц., Чжан И, Гао Дж, Тэн Ф, Тиан В. и др. Висфатин стимулирует пролиферацию клеток рака эндометрия за счет активации сигнальных путей PI3K / Akt и MAPK / ERK1 / 2. Gynecol Oncol. (2016) 143: 168–78. DOI: 10.1016 / j.ygyno.2016.07.109
PubMed Аннотация | CrossRef Полный текст | Google Scholar
60. Зангуэй М., Нурбахш М., Гахремани М.Х., Мешкани Р., Хедри А., Шадбурестан А. и др. Изучение влияния висфатина на фосфорилирование ERalpha (Ser118 и Ser167) и ERE-зависимую транскрипционную активность. EXCLI J. (2018) 17: 516–25. DOI: 10.17179 / excli2018-1299
PubMed Аннотация | CrossRef Полный текст | Google Scholar
61.Захид Х., Суббарамайя К., Айенгар Н.М., Чжоу Х.К., Чен И.С., Бхардвадж П. и др. Лептиновая регуляция оси p53-HIF1alpha / PKM2-ароматаза в стромальных клетках жировой ткани груди: новый механизм связи между ожирением и раком груди. Int J Obes. (2018) 42: 711–20. DOI: 10.1038 / ijo.2017.273
PubMed Аннотация | CrossRef Полный текст | Google Scholar
62. Сон Нью-Йорк, Ли Й.Х., На Х. К., Пэк Дж. Х., Сур Й. Дж. Лептин индуцирует экспрессию SIRT1 через активацию фактора 2, связанного с NF-E2: последствия для канцерогенеза толстой кишки, связанного с ожирением. Biochem Pharmacol. (2018) 153: 282–91. DOI: 10.1016 / j.bcp.2018.02.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
63. Ван П.П., Хэ XY, Ван Р, Ван З., Ван Ю.Г. Высокий уровень лептина — независимый фактор риска рака эндометрия: метаанализ. Cell Physiol Biochem. (2014) 34: 1477–84. DOI: 10.1159 / 000366352
PubMed Аннотация | CrossRef Полный текст | Google Scholar
64. Zhang Y, Liu L, Li C, Ai H. Анализ корреляции между экспрессией лептина и его рецептора (ObR) и клинико-патологией при раке эндометрия. Биомарк рака. (2014) 14: 353–9. DOI: 10.3233 / CBM-140415
PubMed Аннотация | CrossRef Полный текст | Google Scholar
65. Тенвурен И., Дженкс М.З., Рашид Х., Кук К.Л., Мухлеманн Дж. К., Систранк С. и др. Повышенный уровень лептина нарушает полярность эпителия и способствует предраковым изменениям в молочной железе. Онкоген. (2019) 38: 3855–70. DOI: 10.1038 / s41388-019-0687-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
66.Лю Ю, Лв Л, Сяо В, Гун С, Инь Дж, Ван Д. и др. Лептин активирует пути STAT3 и ERK1 / 2 и индуцирует пролиферацию клеток рака эндометрия. J Huazhong Univ Sci Technolog Med Sci. (2011) 31: 365. DOI: 10.1007 / s11596-011-0382-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
67. Bienkiewicz J, Romanowicz H, Malinowski A, Smolarz B. Ассоциация однонуклеотидного полиморфизма — 2548 G / A (rs12112075) гена лептина с раком эндометрия и лейомиомами матки. Eur J Obstet Gynecol Reprod Biol. (2017) 218: 113–8. DOI: 10.1016 / j.ejogrb.2017.09.022
PubMed Аннотация | CrossRef Полный текст | Google Scholar
68. Ван Т., Фарманн Дж. Ф., Ли Х., Ли Й. Дж., Трипати С. К., Юэ С. и др. Бета-окисление жирных кислот, регулируемое JAK / STAT3, имеет решающее значение для самообновления стволовых клеток рака груди и устойчивости к химиотерапии. Cell Metab. (2018) 27: 136–50 e5. DOI: 10.1016 / j.cmet.2017.11.001
CrossRef Полный текст | Google Scholar
69.Цимбалук-Плоска А, Чудецка-Глаз А, Ягодзинская А, Пиус-Садовска Э, Сомпольска-Речула А, Махалински Б и др. Оценка биологически активных веществ, способствующих развитию рака эндометрия или защищающих от него. Onco Targets Ther. (2018) 11: 1363–72. DOI: 10.2147 / OTT.S155942
PubMed Аннотация | CrossRef Полный текст | Google Scholar
70. Дибаба Д.Т., Джадд С.Е., Гилкрист С.К., Кушман М., Пису М., Саффорд М. и др. Связь между ожирением и биомаркерами воспаления и метаболизма со смертностью от рака в проспективном когортном исследовании. Метаболизм. (2019) 94: 69–76. DOI: 10.1016 / j.metabol.2019.01.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
71. Чу Й, Ван И, Пэн В., Сюй Л., Лю М., Ли Дж. И др. Активация STAT3 IL-6 из стволовых клеток, полученных из жировой ткани, способствует пролиферации и метастазированию карциномы эндометрия. Biochem Biophys Res Commun. (2018) 500: 626–31. DOI: 10.1016 / j.bbrc.2018.04.121
PubMed Аннотация | CrossRef Полный текст | Google Scholar
73.He YY, Cai B, Yang YX, Liu XL, Wan XP. Передача сигналов рецептора 30, связанного с эстрогенным G-белком, участвует в регуляции карциномы эндометрия, способствуя пролиферации, потенциалу инвазии и секреции интерлейкина-6 через путь митоген-активируемой протеинкиназы MEK / ERK. Cancer Sci. (2009) 100: 1051–61. DOI: 10.1111 / j.1349-7006.2009.01148.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
74. Че Цюй, Лю BY, Ляо Y, Чжан Х.Дж., Ян Т.Т., Хе YY и др.Активация петли положительной обратной связи с участием ИЛ-6 и ароматазы способствует внутриопухолевому биосинтезу 17-бета-эстрадиола в микросреде карциномы эндометрия. Int J Cancer. (2014) 135: 282–94. DOI: 10.1002 / ijc.28679
PubMed Аннотация | CrossRef Полный текст | Google Scholar
75. Ли Дж. У., Стоун М. Л., Порретт П. М., Томас С. К., Комар Калифорния, Ли Дж. Х. и др. Гепатоциты направляют образование прометастатической ниши в печени. Природа. (2019) 567: 249–52.DOI: 10.1038 / s41586-019-1004-y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
76. Смит Х.О., Стивенс Н.Д., Куоллз С.Р., Флигельман Т., Ван Т., Лин С.Й. и др. Клиническое значение воспалительных цитокинов в первичной культуре клеток рака эндометрия. Mol Oncol. (2013) 7: 41–54. DOI: 10.1016 / j.molonc.2012.07.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
77. Ku SC, Ho PS, Tseng YT, Yeh TC, Cheng SL, Liang CS.Канцерогенез, связанный с бензодиазепином: основное внимание уделяется изменениям биомаркеров рака, связанным с лоразепамом, у лиц с избыточным весом. Психиатрическое расследование. (2018) 15: 900–6. DOI: 10.30773 / pi.2018.05.02.1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
78. Ахмад Р., Аль-Руб А., Кочумон С., Актер Н., Томас Р., Кумари М. и др. Синергизм между пальмитатом и TNF-альфа для продукции CCL2 зависит от пути TRIF / IRF3: последствия для метаболического воспаления. J Immunol. (2018) 200: 3599–611. DOI: 10.4049 / jimmunol.1701552
PubMed Аннотация | CrossRef Полный текст | Google Scholar
79. Доссус Л., Беккер С., Ринальди С., Луканова А., Тьоннеланд А., Олсен А. и др. Фактор некроза опухоли (TNF) -альфа, растворимые рецепторы TNF и риск рака эндометрия: исследование EPIC. Int J Cancer. (2011) 129: 2032–7. DOI: 10.1002 / ijc.25840
PubMed Аннотация | CrossRef Полный текст | Google Scholar
80. Алипурфард I, Датукишвили Н., Микеладзе Д.Подавление TNF-альфа изменяет субстрат рецептора инсулина 1 (IRS-1) в метаболической передаче сигналов диабетических инсулинорезистентных гепатоцитов. Mediat Inflamm. (2019) 2019: 3560819. DOI: 10.1155 / 2019/3560819
PubMed Аннотация | CrossRef Полный текст | Google Scholar
81. Мартинес-Чакон Г., Браун К.А., Доканто М.М., Кумар Х., Салминен С., Сааринен Н. и др. IL-10 подавляет TNF-альфа-индуцированную экспрессию гена ароматазы человека в жировой ткани молочной железы. FASEB J. (2018) 32: 3361–70.DOI: 10.1096 / fj.201700938RRR
PubMed Аннотация | CrossRef Полный текст | Google Scholar
82. Вусден К.А., Лундквист Т., Попович Б., Найман Б., Каррутерс А.М., Ньютон П. и др. Открытие и характеристика антитела, которое избирательно модулирует ингибирующую активность ингибитора-1 активатора плазминогена. Научный доклад (2019) 9: 1605. DOI: 10.2210 / pdb6i8s / PDB
PubMed Аннотация | CrossRef Полный текст | Google Scholar
83. Штайнер Э., Поллоу К., Хазенклевер Д., Шорманн В., Гермес М., Шмидт М. и др.Роль активатора плазминогена урокиназного типа (uPA) и ингибитора активатора плазминогена типа 1 (PAI-1) в прогнозе рака эндометрия. Gynecol Oncol. (2008) 108: 569–76. DOI: 10.1016 / j.ygyno.2007.11.025
PubMed Аннотация | CrossRef Полный текст | Google Scholar
84. Polusani SR, Huang YW, Huang G, Chen CW, Wang CM, Lin LL, et al. Адипокины нарушают регуляцию клеточной коммуникации посредством эпигенетической репрессии локусов щелевых контактов при раке эндометрия с ожирением. Cancer Res. (2019) 79: 196–208. DOI: 10.1158 / 0008-5472.CAN-18-1615
PubMed Аннотация | CrossRef Полный текст | Google Scholar
86. Дили-Конрайт С.М., Парментье Дж. Х., Сами Н., Ли К., Спайсер Д., Мак В. Дж. И др. Воспаление жировой ткани у выживших после рака груди: эффекты 16-недельной комбинированной аэробной тренировки и тренировки с отягощениями. Лечение рака груди. (2018) 168: 147–57. DOI: 10.1007 / s10549-017-4576-y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
87.Соусилло, округ Колумбия, Герриетс В.А., Шенг Дж., Ратмелл Дж. К., Макивер, штат Нью-Джерси. Лептин метаболически лицензирует Т-клетки для активации, чтобы связать питание и иммунитет. J Immunol. (2014) 192: 136–44. DOI: 10.4049 / jimmunol.1301158
PubMed Аннотация | CrossRef Полный текст | Google Scholar
88. Колб Р., Клуз П., Тан З. В., Борчердинг Н., Борман Н., Вишвакарма А. и др. Воспаление, связанное с ожирением, способствует ангиогенезу и раку груди через ангиопоэтин-подобный 4. Онкоген . (2019) 38: 2351–63.DOI: 10.1038 / s41388-018-0592-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
90. Нэгл С.М., Кросби Э.Дж., Бренд А, Обермэр А., Оелер М.К., Куинн М. и др. Связь между диабетом, сопутствующими заболеваниями, индексом массы тела и смертностью от всех причин и от конкретных причин среди женщин с раком эндометрия. Gynecol Oncol. (2018) 150: 99–105. DOI: 10.1016 / j.ygyno.2018.04.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
92.Гу CJ, Xie F, Zhang B, Yang HL, Cheng J, He YY и др. Высокий уровень глюкозы способствует эпителиально-мезенхимальному переходу клеток рака эндометрия матки за счет увеличения ER / GLUT4-опосредованной секреции VEGF. Cell Physiol Biochem. (2018) 50: 706–20. DOI: 10.1159 / 000494237
PubMed Аннотация | CrossRef Полный текст | Google Scholar
93. Валлбиллич Дж. Дж., Джосюла С., Сайни У., Зингарелли Р.А., Дораяппан К.Д., Райли М.К. и др. Высокая глюкозо-опосредованная активация STAT3 при раке эндометрия ингибируется метформином: терапевтические последствия для рака эндометрия. PLoS ONE. (2017) 12: e0170318. DOI: 10.1371 / journal.pone.0170318
PubMed Аннотация | CrossRef Полный текст | Google Scholar
95. Чжун XY, Юань XM, Xu YY, Yin M, Yan WW, Zou SW и др. CARM1 метилирует GAPDH для регулирования метаболизма глюкозы и подавляется при раке печени. Cell Rep. (2018) 24: 3207–23. DOI: 10.1016 / j.celrep.2018.08.066
PubMed Аннотация | CrossRef Полный текст | Google Scholar
96. Wei Y, Wang D, Jin F, Bian Z, Li L, Liang H, et al.Пируваткиназа типа M2 способствует высвобождению экзосом опухолевых клеток посредством фосфорилирования белка 23, связанного с синаптосомами. Nat Commun. (2017) 8: 14041. DOI: 10.1038 / ncomms14041
PubMed Аннотация | CrossRef Полный текст | Google Scholar
97. Бон Т., Рапп С., Лютер Н., Кляйн М., Брюль Т.Дж., Кодзима Н. и др. Иммуноэвазия опухолей через ацидоз-зависимую индукцию регуляторных макрофагов, ассоциированных с опухолью. Nat Immunol. (2018) 19: 1319–29. DOI: 10.1038 / s41590-018-0226-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
98.Гу С, Ни Т, Ван Дж, Лю И, Фан Кью, Ван И и др. Блокада CD47 подавляет прогрессирование опухоли, способствуя фагоцитозу опухолевых клеток макрофагами поляризованными M2 при раке эндометрия. J Immunol Res. (2018) 2018: 6156757. DOI: 10.1155 / 2018/6156757
PubMed Аннотация | CrossRef Полный текст | Google Scholar
99. Латиф А., Чедвик А.Л., Китсон С.Дж., Грегсон Х.Дж., Сивалингам В.Н., Болтон Дж. И др. Транспортер монокарбоксилата 1 (MCT1) является независимым прогностическим биомаркером рака эндометрия. BMC Clin Pathol. (2017) 17:27. DOI: 10.1186 / s12907-017-0067-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
100. Лю Н, Ло Дж, Куанг Д., Сюй С., Дуань И, Ся И и др. Лактат подавляет экспрессию ATP6V0d2 в связанных с опухолью макрофагах, способствуя HIF-2альфа-опосредованному прогрессированию опухоли. J Clin Invest. (2019) 129: 631–46. DOI: 10.1172 / JCI123027
PubMed Аннотация | CrossRef Полный текст | Google Scholar
101. Friedenreich CM, Langley AR, Speidel TP, Lau DC, Courneya KS, Csizmadi I, et al.Исследование маркеров инсулинорезистентности и риска рака эндометрия методом случай-контроль. Endocr Relat Cancer. (2012) 19: 785–92. DOI: 10.1530 / ERC-12-0211
PubMed Аннотация | CrossRef Полный текст | Google Scholar
102. Кабат Г.К., Ким М.Ю., Лейн Д.С., Заславский О., Хо ГЙФ, Луо Дж. И др. Глюкоза и инсулин в сыворотке и риск рака груди, эндометрия и яичников у женщин в постменопаузе. Eur J Cancer Пред. (2018) 27: 261–8. DOI: 10.1097 / CEJ.0000000000000435
PubMed Аннотация | CrossRef Полный текст | Google Scholar
104.Джоелин-Прайс А.С., Стивенс Дж.А., Чжан Дж., Бэкес Ф.Дж., Кон Д.Е., Суарес А.А. Рак эндометрия Экспрессия рецептора инсулиноподобного фактора роста 1 (IGF1R) увеличивается с индексом массы тела и связана с патологической степенью и прогнозом. Биомаркеры эпидемиологии рака Пред. (2016) 25: 438–45. DOI: 10.1158 / 1055-9965.EPI-15-1145
PubMed Аннотация | CrossRef Полный текст | Google Scholar
105. Ван Ц., Су К., Чжан И, Чжан В., Чжао Ц., Чу Д. и др. Сигналы, опосредованные IR-A / IGF-1R, способствуют эпителиально-мезенхимальному переходу клеток карциномы эндометрия путем активации путей PI3K / AKT и ERK. Cancer Biol Ther. (2019) 20: 295–306. DOI: 10.1080 / 15384047.2018.1529096
PubMed Аннотация | CrossRef Полный текст | Google Scholar
106. Zhang H, Fagan DH, Zeng X, Freeman KT, Sachdev D, Yee D. Ингибирование пролиферации и метастазирования раковых клеток путем подавления рецепторов инсулина. Онкоген. (2010) 29: 2517–27. DOI: 10.1038 / onc.2010.17
PubMed Аннотация | CrossRef Полный текст | Google Scholar
107. Крайсер Дж. Р., Юги К., Хираяма А., Фазакерли Д. Д., Квек Л. Е., Скалцо Р. и др.Динамическая метаболомика показывает, что инсулин настраивает адипоциты для метаболизма глюкозы. Cell Rep. (2017) 21: 3536–47. DOI: 10.1016 / j.celrep.2017.11.085
PubMed Аннотация | CrossRef Полный текст | Google Scholar
108. Zhou R, Yang Y, Lu Q, Wang J, Miao Y, Wang S, et al. Факторы прогноза онкологических и репродуктивных исходов при лечении сложной атипичной гиперплазии и рака эндометрия низкой степени злокачественности с использованием перорального прогестина у пациентов из Китая с сохранением фертильности. Gynecol Oncol. (2015) 139: 424–8. DOI: 10.1016 / j.ygyno.2015.09.078
PubMed Аннотация | CrossRef Полный текст | Google Scholar
109. Хейрандиш М., Махбуби Х., Язданпараст М., Камаль В., Камал М.А. Противораковые эффекты метформина: недавние доказательства его роли в профилактике и лечении рака. Curr Drug Metab. (2018) 19: 793–7. DOI: 10.2174 / 138
19666180416161846
PubMed Аннотация | CrossRef Полный текст | Google Scholar
110.Кантрелл Л.А., Чжоу К., Мендивил А., Маллой К.М., Гериг П.А., Бэ-Джамп В.Л. Метформин является мощным ингибитором пролиферации клеток рака эндометрия, что имеет значение для новой стратегии лечения. Gynecol Oncol. (2010) 116: 92–8. DOI: 10.1016 / j.ygyno.2009.09.024
PubMed Аннотация | CrossRef Полный текст | Google Scholar
111. Невадунский Н.С., Ван Арсдейл А., Стриклер Х.Д., Моадель А., Каур Г., Фример М. и др. Использование метформина и выживаемость при раке эндометрия. Gynecol Oncol. (2014) 132: 236–40. DOI: 10.1016 / j.ygyno.2013.10.026
PubMed Аннотация | CrossRef Полный текст | Google Scholar
112. Китсон С.Дж., Маскелл З., Сивалингам В.Н., Аллен Дж.Л., Али С., Бернс С. и др. Предоперационное исследование метформина при злокачественных новообразованиях матки (ПРЕМИУМ): многоцентровое рандомизированное двойное слепое плацебо-контролируемое исследование III фазы. Clin Cancer Res. (2019) 25: 2424–32. DOI: 10.1158 / 1078-0432.CCR-18-3339
PubMed Аннотация | CrossRef Полный текст | Google Scholar
113.Артур Р., Моллер Х., Гармо Х., Хаггстром С., Холмберг Л., Статтин П. и др. Глюкоза, триглицериды и холестерин в сыворотке в отношении смерти от рака простаты в шведском исследовании AMORIS. Контроль причин рака. (2019) 30: 195–206. DOI: 10.1007 / s10552-018-1093-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
114. Артур Р.С., Кабат Г.К., Ким М.Ю., Уайлд Р.А., Шадьяб А.Х., Вактавски-Венде Дж. И др. Метаболический синдром и риск рака эндометрия у женщин в постменопаузе: проспективное исследование. Контроль причин рака. (2019) 30: 355–63. DOI: 10.1007 / s10552-019-01139-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
115. Брюнинг П.Ф., Бонфрер Дж. М.. Концентрации свободных жирных кислот коррелировали с доступной фракцией эстрадиола в плазме крови человека. Cancer Res. (1986) 46: 2606–9.
PubMed Аннотация | Google Scholar
116. Шмандт Р. Э., Иглесиас Д. А., Ко Н. Н., Лу К. Х. Понимание ожирения и риска рака эндометрия: возможности для профилактики. Am J Obstet Gynecol. (2011) 205: 518–25. DOI: 10.1016 / j.ajog.2011.05.042
PubMed Аннотация | CrossRef Полный текст | Google Scholar
117. Гибсон Д.А., Коллинз Ф., Казинс Флорида, Эсналь Суфиаурре А, Сондерс ПТК. Влияние 27-гидроксихолестерина на пролиферацию рака эндометрия. Endocr Relat Cancer. (2018) 25: 381–91. DOI: 10.1530 / ERC-17-0449
PubMed Аннотация | CrossRef Полный текст | Google Scholar
118. Madak-Erdogan Z, Band S, Zhao YC, Smith BP, Kulkoyluoglu-Cotul E, Zuo Q, et al.Свободные жирные кислоты изменяют метаболизм рака молочной железы, связанного с ожирением, через рецептор эстрогена и передачу сигналов mTOR. Cancer Res. (2019) 79: 2494–510. DOI: 10.1158 / 0008-5472.CAN-18-2849
PubMed Аннотация | CrossRef Полный текст | Google Scholar
119. Хао Дж, Чжан И, Янь Х, Янь Ф, Сунь Й, Цзэн Дж и др. Циркулирующий белок, связывающий жирные кислоты жировой ткани, является новым звеном, лежащим в основе связанного с ожирением развития опухоли груди / молочной железы. Cell Metab. (2018) 28: 689–705 e5.DOI: 10.1016 / j.cmet.2018.07.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
120. Бринтон Л.А., Сакода Л.С., Фредериксен К., Шерман М.Э., Кьяер С.К., Граубард Б.И. и др. Связь опухолей матки и яичников с уже существующими хроническими состояниями. Gynecol Oncol. (2007) 107: 487–94. DOI: 10.1016 / j.ygyno.2007.08.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
121. Зеебахер В., Хофштеттер Г., Полтерауер С., Рейнталлер А., Гримм С., Швамейс Р. и др.Влияет ли тиреотропный гормон на прогноз больных раком эндометрия? Многоцентровое исследование. Br J Рак. (2013) 109: 215–8. DOI: 10.1038 / bjc.2013.282
PubMed Аннотация | CrossRef Полный текст | Google Scholar
122. Шинков А., Борисова А.М., Ковачева Р., Атанасова И., Влахов Дж., Даковска Л. Распространенность метаболического синдрома увеличивается за счет квартилей тиреотропного гормона в популяционной выборке эутиреоидных субъектов. Arq Bras Endocrinol Metabol. (2014) 58: 926–32. DOI: 10.1590 / 0004-2730000003538
PubMed Аннотация | CrossRef Полный текст | Google Scholar
125. МакКинтош М.Л., Дербишир А.Э., Маквей Р.Дж., Болтон Дж., Никко-Амири М., Хиггинс С.Л. и др. Влияние ожирения и бариатрической хирургии на циркулирующие и тканевые биомаркеры риска рака эндометрия. Int J Cancer. (2019) 144: 641–50. DOI: 10.1002 / ijc.31913
PubMed Аннотация | CrossRef Полный текст | Google Scholar
126.Линьков Ф., Гоунур С.Л., Ма Т, Сюй З., Эдвардс Р.П., Локшин А.Е. и др. Изменения биомаркеров риска воспалительного рака эндометрия у лиц, перенесших хирургическую потерю веса. Gynecol Oncol. (2017) 147: 133–8. DOI: 10.1016 / j.ygyno.2017.07.144
PubMed Аннотация | CrossRef Полный текст | Google Scholar
127. Буш Э.Л., Кроус-Боу М., Прескотт Дж., Даунинг М.Дж., Рознер Б.А., Муттер Г.Л. и др. Адипонектин, лептин и рецепторы инсулинового пути как маркеры подтипов рака эндометрия. Horm Cancer. (2018) 9: 33–9. DOI: 10.1007 / s12672-017-0318-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
128. Китсон С.Дж., Эванс Д.Г., Кросби Э.Дж. Выявление женщин из группы высокого риска для стратегий профилактики рака эндометрия: предложение модели прогнозирования риска рака эндометрия. Cancer Prev Res. (2017) 10: 1–13. DOI: 10.1158 / 1940-6207.CAPR-16-0224
PubMed Аннотация | CrossRef Полный текст | Google Scholar
129. Fortner RT, Husing A, Kuhn T., Konar M, Overvad K, Tjonneland A, et al.Прогнозирование риска рака эндометрия, включая сывороточные биомаркеры: результаты когорты EPIC. Int J Cancer. (2017) 140: 1317–23. DOI: 10.1002 / ijc.30560
PubMed Аннотация | CrossRef Полный текст | Google Scholar
130. Линь И, Чжоу Дж, Цао Л., Сюй Q, Хао Дж, Чжао Л. и др. Кальций в сыворотке крови — новый параметр для оценки метаболического синдрома при карциноме эндометрия. J Gynecol Oncol. (2019) 30: e12. DOI: 10.3802 / jgo.2019.30.e12
PubMed Аннотация | CrossRef Полный текст | Google Scholar
1 | Леманн Б.Д., Бауэр Дж.А., Чен Х, Сандерс М.Э., Чакраварти А.Б., Шир Й. и Питенпол Дж.А.: Идентификация человека. подтипы тройного отрицательного рака молочной железы и доклинические модели для подбор таргетной терапии.J Clin Invest. 121: 2750–2767. 2011.PubMed / NCBI |
2 | Леманн Б.Д., Йованович Б., Чен Х, Эстрада М.В., Джонсон К.Н., Шир Й., Моисей Х.Л., Сандерс М.Э. и Питенпол Дж.А.: Уточнение молекулярных подтипов рака молочной железы с тройным отрицательным результатом: Значение для выбора неоадъювантной химиотерапии. PLoS One. 11: e01573682016.PubMed / NCBI |
3 | Джулиано А.Е., Коннолли Д.Л., Эдж С.Б., Mittendorf EA, Rugo HS, Solin LJ, Weaver DL, Winchester DJ и Хортобаджи Г.Н.: Основные изменения в американском Джойнте — рак груди Комитет по раку, восьмое издание руководства по стадированию рака.CA Рак J Clin. 67: 290–303. 2017.PubMed / NCBI |
4 | Вольф AC, Хаммонд MEH, Эллисон К.Х., Харви BE, Мангу ПБ, Бартлетт Дж.М.С., Билоус М., Эллис И.О., Фитцгиббонс П., Hanna W и др.: Тестирование рецептора 2 эпидермального фактора роста человека в рак груди: Американское общество клинической онкологии / Колледж Обновление рекомендаций по клинической практике американских патологов. J Clin Oncol. 36: 2105–2122. 2018.PubMed / NCBI |
5 | Эллисон К. Х., Хаммонд М. Е., Доусет М., МакКернин С.Е., Кэри Л.А., Фицгиббонс П.Л., Хейс Д.Ф., Лакани С.Р., Чавес-МакГрегор М., Перлмуттер Дж. И др.: Эстроген и прогестерон. тестирование рецепторов при раке груди: обновление рекомендаций ASCO / CAP.J Clin Oncol. 38: 1346–1366. 2020.PubMed / NCBI |
6 | Yam C, Mani SA и Moulder SL: нацеливание молекулярные подтипы тройного отрицательного рака груди: Понимание разнообразия для прогресса в этой области. Онколог. 22: 1086–1093. 2017.PubMed / NCBI |
7 | Бонотто М, Герратана Л, Полетто Е, Дриол P, Giangreco M, Russo S, Minisini AM, Andreetta C, Mansutti M, Пиза FE и др.: Оценка результатов при метастатическом раке молочной железы: Выводы из реального сценария.Онколог. 19: 608–615. 2014.PubMed / NCBI |
8 | Колер Б.А., Шерман Р.Л., Ховладер Н., Джемаль А, Райерсон А.Б., Генри К.А., Боско Ф.П., Кронин К.А., Лейк А, Нун А.М. и др. al: Годовой отчет для страны о статусе онкологических заболеваний, 1975-2011 гг. с указанием заболеваемости подтипами рака груди по расе / этнической принадлежности, бедность и состояние. J Natl Cancer Inst. 107: djv0482015. |
9 | О’Шонесси Дж., Шварцберг Л., Дансо М. А., Миллер К.Д., Руго Х.С., Нойбауэр М., Роберт Н., Хеллерстедт Б., Салех М., Richards P и др.: Исследование фазы III комбинации инициариба плюс гемцитабин и карбоплатин по сравнению с гемцитабином и карбоплатином у пациентов с метастатический тройной отрицательный рак груди.J Clin Oncol. 32: 3840–3847. 2014.PubMed / NCBI |
10 | Caswell-Jin JL, Plevritis SK, Tian L, Cadham CJ, Xu C, Stout NK, Sledge GW, Mandelblatt JS и Kurian AW: Изменение выживаемости при метастатическом раке молочной железы на фоне лечения достижения: метаанализ и систематический обзор. JNCI Cancer Spectr. 2: pky0622018. |
11 | Плеврит СК, Муньос Д., Куриан А.В., Стаут NK, Alagoz O, Near AM, Lee SJ, van den Broek JJ, Huang X, Schechter CB, et al: Ассоциация скрининга и лечения груди смертность от рака по молекулярным подтипам у женщин в США, 2000-2012 гг.ДЖАМА. 319: 154–164. 2018. Просмотр статьи: Google Scholar: PubMed / NCBI |
12 | Стэнтон С.Е., Адамс С. и Дисис М.Л .: Различия в частоте и масштабах инфильтрации опухоли лимфоциты в подтипах рака груди: систематический обзор. JAMA Онкол. 2: 1354–1360. 2016. Просмотреть статью: Google Scholar: PubMed / NCBI |
13 | Сафонов А., Цзян Т., Бьянчини Дж., Дьерфи B, Karn T, Hatzis C и Pusztai L: экспрессия иммунных генов связаны с геномными аберрациями при раке груди.Cancer Res. 77: 3317–3324. 2017. Просмотр статьи: Google Scholar: PubMed / NCBI |
14 | Шмид П., Адамс С., Руго Х.С., Шнейвайс А., Барриос С.Х., Ивата Х., Диерас В., Хегг Р., Им С.А., Шоу Райт Г. и др.: Атезолизумаб и наб-паклитаксел при прогрессирующей тройной отрицательной груди рак. N Engl J Med. 379: 2108–2121. 2018. Просмотр статьи: Google Scholar: PubMed / NCBI |
15 | Шмид П., Руго Х.С., Адамс С., Шнейвайс А, Барриос С.К., Ивата Х., Диерас В., Хеншель В., Молинеро Л., Чуй С.И. и др. др.: атезолизумаб плюс наб-паклитаксел в качестве лечения первой линии при неоперабельная, местнораспространенная или метастатическая тройная отрицательная грудь рак (IMpassion130): обновленные результаты эффективности рандомизированного, двойное слепое плацебо-контролируемое исследование фазы 3.Ланцет Онкол. 21: 44–59. 2020. Просмотр статьи: Google Scholar |
16 | Дадли Дж. К., Лин М. Т., Ле Д. Т. и Эшлеман Дж. Р.: Нестабильность микросателлитов как биомаркер блокады PD-1. Clin Cancer Res. 22: 813–820. 2016. Просмотреть статью: Google Scholar: PubMed / NCBI |
17 | Бонневиль Р., Крук М.А., Каутто Е.А., Мия Дж., Wing MR, Chen HZ, Reeser JW, Yu L и Roychowdhury S: Пейзаж микросателлитная нестабильность при 39 типах рака.JCO Precis Онкол. 2017: ПО.17.00073. 2017.PubMed / NCBI |
18 | Курата К., Кубо М., Мори Х., Кавадзи Х., Мотояма Ю., Куроки Л., Ямада М., Канеширо К., Кай М. и Накамура М.: Нестабильность микросателлитов при тройном отрицательном раке молочной железы. В: Материалы симпозиума по раку груди в Сан-Антонио 2018 г. Рак Res. 79 (Приложение 4): Аннотация № P1-06-11. 2019. |
19 | Брайант Х.Э., Шульц Н., Томас HD, Паркер К.М., Флауэр Д., Лопес Э., Кайл С., Меут М., Куртин Нью-Джерси и Хелледей Т.: Специфическое уничтожение BRCA2-дефицитных опухолей ингибиторами поли (АДФ-рибоза) полимераза.Природа. 434: 913–917. 2005. Просмотр статьи: Google Scholar: PubMed / NCBI |
20 | Фермер Х., МакКейб Н., лорд С.Дж., Тутт А.Н., Джонсон Д.А., Ричардсон ТБ, Сантароза М., Диллон К.Дж., Хиксон И., Knights C и др.: Нацеливание на дефект репарации ДНК у мутанта BRCA клетки как терапевтическая стратегия. Природа. 434: 917–921. 2005 г. Просмотр статьи: Google Scholar: PubMed / NCBI |
21 | Робсон М., Им С.А., Сенкус Э., Сюй Б., Домчек С.М., Масуда Н., Делалог С., Ли В., Тунг Н., Армстронг А. и др.: Олапариб при метастатическом раке груди у пациентов с зародышевой линией Мутация BRCA.N Engl J Med. 377: 523–533. 2017. Просмотр статьи: Google Scholar: PubMed / NCBI |
22 | Робсон М.Э., Тунг Н, Конте П, Им С.А., Сенкус Э, Сюй Б., Масуда Н., Делалог С., Ли В., Армстронг А. и др.: OlympiAD окончательные результаты общей выживаемости и переносимости: олапариб по сравнению с химиотерапевтическое лечение по выбору врача у пациентов с мутация BRCA зародышевой линии и HER2-отрицательный метастатический рак молочной железы. Энн Онкол. 30: 558–566. 2019. Просмотр статьи: Google Scholar: PubMed / NCBI |
23 | Литтон Дж. К., Руго Х. С., Эттл Дж., Гурвиц С.А., Гонсалвеш А., Ли К. Х., Ференбахер Л., Йерушалми Р., Мина Л. А., Мартин M, et al: Талазопариб у пациентов с распространенным раком молочной железы и мутация BRCA зародышевой линии.N Engl J Med. 379: 753–763. 2018. Просмотр статьи: Google Scholar: PubMed / NCBI |
24 | Денкерт К., фон Минквиц Г., Дарб-Исфахани С., Ледерер Б., Хеппнер Б.И., Вебер К.Э., Будцис Дж., Хубер Дж., Клаушен F, Furlanetto J, et al: Лимфоциты, инфильтрирующие опухоль и прогноз при различных подтипах рака груди: объединенный анализ 3771 пациента, получавшего неоадъювантную терапию. Ланцет Онкол. 19: 40–50. 2018. Просмотр статьи: Google Scholar |
25 | Хида А.И., Ватанабэ Т., Сагара Й., Кашиваба M, Sagara Y, Aogi K, Ohi Y и Tanimoto A: диффузное распределение лимфоциты, инфильтрирующие опухоль, являются маркером для лучшего прогноза и химиотерапевтический эффект при тройном отрицательном раке груди.Грудь Лечение рака Res. 178: 283–294. 2019. Просмотр статьи: Google Scholar: PubMed / NCBI |
26 | Лой С., Друбай Д., Адамс С., Прунери Дж., Francis PA, Lacroix-Triki M, Joensuu H, Dieci MV, Badve S, Demaria S и др.: Лимфоциты, инфильтрирующие опухоль, и прогноз: объединенный индивидуальный анализ пациента тройной отрицательной молочной железы на ранней стадии раки. J Clin Oncol. 37: 559–569. 2019. Просмотр статьи: Google Scholar: PubMed / NCBI |
27 | Галон Дж и Бруни Д: подходы к лечению иммунные горячие, измененные и холодные опухоли с комбинацией иммунотерапия.Nat Rev Drug Discov. 18: 197–218. 2019. Просмотр статьи: Google Scholar: PubMed / NCBI |
28 | Адамс С., Гатти-Мэйс М.Э., Калинский К., Корде Лос-Анджелес, Шарон Э., Амири-Кордестани Л., Медведь Х., МакАртур Х. Л., Фрэнк Э., Перлмуттер Дж. И др.: Современное состояние иммунотерапии груди. рак: обзор. JAMA Oncol. 11–2019 апреля. Epub в преддверии печати. Просмотр статьи: Google Scholar |
29 | Шмид П., Сальгадо Р., Парк YH, Муньос-Кусело Э., Ким С.Б., Сон Дж., Им С.А., Фукакис Т., Куэммел С., Дент Р. и др.: Пембролизумаб плюс химиотерапия как неоадъювант лечение тройного отрицательного рака молочной железы с высоким риском на ранней стадии: Результаты открытого, многоконтактного этапа 1b KEYNOTE-173 изучение.Энн Онкол. 31: 569–581. 2020. Просмотр статьи: Google Scholar: PubMed / NCBI |
30 | Нанда Р., Лю М.С., Яу С., Шацкий Р., Пуштай Л., Уоллес А., Чиен А.Дж., Фореро-Торрес А., Эллис Э., Хан Х и др.: Влияние пембролизумаба в сочетании с неоадъювантной химиотерапией на патологические полный ответ у женщин с раком груди на ранней стадии: анализ продолжающегося фазы 2 адаптивно рандомизированного исследования I-SPY2. JAMA Oncol. 6: 1–9. 2020. Просмотр статьи: Google Scholar |
31 | Шмид П., Кортес Дж., Пуштаи Л., МакАртур Х., Кюммель С., Берг Дж., Денкерт С., Парк Ю. Х., Хуэй Р., Харбек Н. и др.: Пембролизумаб при раннем тройном отрицательном раке молочной железы.N Engl J Med. 382: 810–821. 2020. Просмотр статьи: Google Scholar: PubMed / NCBI |
32 | Джанни Л., Хуанг С.С., Эгле Д., Бермеджо Б. Zamagni C, Thill M, Anton A, Zambelli S, Bianchini G, Russo S и Цируелос Э: Патологический полный ответ (pCR) на неоаджвонт лечение с атезолизумабом или без него при тройном отрицательном результате, в начале местно-распространенный рак груди высокого риска. NeoTRIPaPDL1 Микеланджело рандомизированное исследование. В: Труды San 2019 Симпозиум Антонио по раку груди.Cancer Res. 80 (Приложение 4): Аннотация № GS3-04. 2020. |
33 | Миттендорф Э, Барриос СН, Харбек Н, Майлз D, Saji S, Zhang H, Duc AN, Rafii S и Lai C: IMpassion031: A исследование фазы III, сравнивающее неоадъювантный атезолизумаб и плацебо в комбинация с химиотерапией на основе наб-паклитаксела на ранней стадии тройной отрицательный рак груди (TNBC). В: Материалы 2017 г. Симпозиум по раку груди в Сан-Антонио. Cancer Res. 78 (Дополнение 4): Реферат № ОТ2-07-03.2018. |
34 | Cortes J, Cescon DW, Rugo HS, Nowecki Z, Им С.А., Юсоф М.М., Галлардо К., Липатов О., Барриос С.Х., Ольгадо Э. и др. al: KEYNOTE-355: Рандомизированное двойное слепое исследование III фазы пембролизумаб + химиотерапия по сравнению с плацебо + химиотерапия для ранее не леченный местно рецидивирующий неоперабельный или метастатический тройной отрицательный рак груди. J Clin Oncol. 38 (Приложение 15): S1000. 2020. Просмотр статьи: Google Scholar |
35 | Voorwerk L, Slagter M, Horlings HM, Сикорска К., ван де Вейвер К.К., де Маакер М., Недерлоф И., Клюин RJC, Уоррен С., Онг С. и др.: Стратегии иммунной индукции в метастатике тройной отрицательный рак молочной железы для повышения чувствительности к PD-1 блокада: испытание TONIC.Nat Med. 25: 920–928. 2019. Просмотр статьи: Google Scholar: PubMed / NCBI |
36 | Аллард Б., Лонги М.С., Робсон СК и Стэгг J: эктонуклеотидазы CD39 и CD73: новый ингибитор контрольной точки цели. Immunol Rev.276: 121–144. 2017. Просмотр статьи: Google Scholar: PubMed / NCBI |
37 | Галамфарса Г., Каземи М. Х., Рауфи Мохсени С., Масджеди А., Ходжат-Фарсанги М., Азизи Г., Юсефи М. и Джадиди-Ниараг F: CD73 как потенциальная возможность для рака иммунотерапия.Эксперт считает, что цели. 23: 127–142. 2019. Просмотр статьи: Google Scholar |
38 | Duhant X, Schandené L, Bruyns C, Gonzalez NS, Goldman M, Boeynaems JM и Communi D: внеклеточный аденин нуклеотиды подавляют активацию CD4 + Т-лимфоцитов человека. J Иммунол. 169: 15–21. 2002. Просмотр статьи: Google Scholar: PubMed / NCBI |
39 | Аллард Б., Бивис П.А., Дарси П.К. и Стэг Дж.: Иммуносупрессивная активность аденозина при раке.Curr Opin Pharmacol. 29: 7–16. 2016. Просмотреть статью: Google Scholar: PubMed / NCBI |
40 | Охта А. Контрольная точка метаболического иммунитета: Аденозин в микросреде опухоли. Фронт Иммунол. 7: 10 . Просмотр статьи: Google Scholar: PubMed / NCBI |
41 | Buisseret L, Pommey S, Allard B, Garaud S, Bergeron M, Cousineau I, Ameye L, Bareche Y, Paesmans M, Crown JPA, и др.: Клиническое значение CD73 в трижды отрицательной груди. рак: Мультиплексный анализ клинического исследования фазы III.Аня Онкол. 29: 1056–1062. 2018. |
42 | Пудровый Дж, Спира А, Гутьеррес Р, ДиРензо D, Udyavar A, Karakunnel JJ, Rieger A, Colabella J, Lai DW и де Соуза П: оценка фазы 1 AB928, нового двойного аденозина антагонист рецепторов, в сочетании с химиотерапией или AB122 (анти-PD-1) у пациентов с запущенными злокачественными новообразованиями. Энн Онкол. 30 (Дополнение 5): v475 – v532. 2019. |
43 | Хартман А.Р., Калдате Р.Р., Зайлер Л.М., Художник Л., Гриер К.Э., Эндсли Р.Р., Гриффин М., Гамильтон С.А., Фрай Калифорния, Зильберман MA и др.: Распространенность мутаций BRCA в невыбранной популяции тройного отрицательного рака груди.Рак. 118: 2787–2795. 2012.PubMed / NCBI |
44 | Okuma HS и Yonemori K: ген BRCA мутации и ингибиторы поли (АДФ-рибозы) полимеразы в тройной отрицательный рак груди. Adv Exp Med Biol. 1026: 271–286. 2017.PubMed / NCBI |
45 | Лорд CJ и Эшворт A: BRCAness пересмотрел. Nat Rev Рак. 16: 110–120. 2016.PubMed / NCBI |
46 | Литтон Дж. К., Скоггинс М. Е., Хесс К. Р., Адрада BE, Мурти Р.К., Дамодаран С., ДеСнайдер С.М., Брюстер А.М., Барсенас СН, Валеро V и др.: Неоадъювантный талазопариб для пациентов с операбельным рак груди с патогенным вариантом зародышевой линии BRCA.J Clin Онкол. 38: 388–394. 2020. |
47 | Лойбл С., О’Шонесси Дж., Унч М., Сиков WM, Руго HS, McKee MD, Huober J, Golshan M, von Minckwitz G, Maag D, et al: Добавление ингибитора PARP велипариба плюс карбоплатин или только карбоплатин к стандартной неоадъювантной химиотерапии при тройной отрицательный рак груди (яркость): рандомизированный, фаза 3 пробный. Ланцет Онкол. 19: 497–509. 2018.PubMed / NCBI |
48 | Цзяо С., Ся В, Ямагути Х, Вэй И, Чен MK, Hsu JM, Hsu JL, Yu WH, Du Y, Lee HH и др.: Ингибитор PARP усиливает экспрессию PD-L1 и усиливает связанный с раком иммуносупрессия.Clin Cancer Res. 23: 3711–3720. 2017.PubMed / NCBI |
49 | Винаяк С, Толани С.М., Шварцберг Л, Mita M, McCann G, Tan AR, Wahner-Hendrickson AE, Forero A, Anders C, Wulf GM и др.: Открытое клиническое испытание комбинации нирапариба с пембролизумабом для лечения запущенных или метастатических тройной отрицательный рак груди. JAMA Oncol. 5: 1132–1140. 2019. |
50 | Domchek S, Postel-Vinay S, Im S, Park YH, Делорд Дж., Итальяно А, Александр Дж., Ю Б., Бастиан С., Кребс М. Г. и др. al: Фаза II исследования олапариба (о) и дурвалумаба (d) (MEDIOLA): Обновленные результаты у пациентов с мутацией BRCA зародышевой линии (gBRCAm) метастатический рак молочной железы (mbc).Энн Онкол. 30 (Дополнение 5): v475 – v532. 2019. |
51 | Пуштаи Л., Хан Х.С., Яу С., Вольф Д., Уоллес А.М., Шацкий Р., Хелстен Т., Бугхи Дж. К., Хаддад Т., Стрингер-Реазор Э., и др.: Дурвалумаб в комбинации с олапарибом и паклитакселом в HER2-отрицательный рак молочной железы стадии II / III высокого риска: результаты испытание I-SPY 2. В: Материалы ежегодного собрания Американская ассоциация исследований рака 2020. Cancer Res. 80 (Доп. 16): Реферат № CT011.2020. |
52 | Митри З.И., Вуки Дж., Кеммер К.А., Савин М.А., Пармар С., Колодзи А.К., Джонсон Б., Уильямс-Белизер Р., Грей Дж. У. и Миллс, Великобритания: исследование II фазы олапариба и дурвалумаба при метастазах. BRCA тройной отрицательный рак молочной железы дикого типа. J Clin Oncol. 37: TPS11112019. |
53 | Руго Х.С., Лломбарт-Кюссак А, Андре Ф, Робсон М.Э., Саджи С., Харбек Н., Шмид П., Сескон Д. В., Ан Дж. С., Нанда Р., et al: KEYLYNK-009: открытое рандомизированное исследование фазы II / III пембролизумаб (пембро) плюс олапариб против пембро плюс химиотерапия после индукции пембро первой линии плюс химиотерапия в пациенты с местно рецидивирующими неоперабельными или метастатическими тройной отрицательный рак груди (TNBC).J Clin Oncol. 38: TPS5962020. |
54 | Maacke H, Opitz S, Jost K, Hamdorf W, Хеннинг В., Крюгер С., Феллер А.С., Лопенс А., Дидрих К., Швингер Е. и Stürzbecher HW: сверхэкспрессия коррелятов Rad51 дикого типа с гистологической классификацией инвазивного протокового рака молочной железы. Int J Рак. 88: 907–913. 2000.PubMed / NCBI |
55 | Мартин Р.В., Орелли Б.Дж., Ямазо М, Минн А.Дж., Takeda S и Bishop DK: повышающая регуляция RAD51 обходит функцию BRCA1 и является общим признаком опухолей молочной железы с дефицитом BRCA1.Рак Res. 67: 9658–9665. 2007. PubMed / NCBI |
56 | Wiegmans AP, Yap PY, Ward A, Lim YC и Ханна К.К .: Различия в экспрессии ключевых генов восстановления повреждений ДНК после того, как эпигенетически индуцированная BRCAness диктует синтетическую летальность с Ингибирование PARP1. Mol Cancer Ther. 14: 2321–2331. 2015.PubMed / NCBI |
57 | Лю И, Бернесс М.Л., Мартин-Тревино Р., Гай Дж., Бай С., Харуака Р., Брукс М.Д., Шан Л., Фокс А., Лютер Т.К. и др.: RAD51 обеспечивает устойчивость раковых стволовых клеток к ингибированию PARP при тройном отрицательном раке груди.Clin Cancer Res. 23: 514–522. 2017. |
58 | Марцио А., Пуччини Дж., Квон Й., Маверакис Н.К., Арбини А., Сун П., Бар-Саги Д. и Пагано М.: The F-Box доменно-зависимая активность EMI1 регулирует чувствительность PARPi в тройной отрицательный рак груди. Mol Cell. 73: 224–237.e6. 2019. |
59 | Тутт А., Стивенс С., Фруер П., Пирс А., Rhee J, So K, Ottesen L, Dean E и Hollingsworth SJ: ФИОЛЕТКА: A рандомизированное исследование фазы II для оценки ингибиторов ответа на повреждение ДНК в сочетании с олапарибом (Ола) vs.Монотерапия Ола у пациентов (пациенты) с метастатическим трижды отрицательным раком молочной железы (TNBC) стратифицированный изменениями в репарации гомологичной рекомбинации (HRR) -связанные гены. J Clin Oncol. 36 (Приложение 15): TPS11122018. |
60 | Хираи Х, Араи Т, Окада М, Нисибата Т, Кобаяси М., Сакаи Н., Имагаки К., Отани Дж., Сакаи Т., Йошизуми Т. и др. al: MK-1775, низкомолекулярный ингибитор Wee1, усиливает противоопухолевую эффективность различных ДНК-повреждающих агентов, в том числе 5-фторурацила.Cancer Biol Ther. 9: 514–522. 2010.PubMed / NCBI |
61 | Питтс TM, Симмонс Д.М., Бэгби С.М., Хартман SJ, Якоб Б.В., Гиттлман Б., Тентлер Джей-Джей, Ситтелли Д., Ормонд Д.Р., Мессерсмит В.А. и др.: Ингибирование Wee1 усиливает противоопухолевый эффект. эффекты капецитабина в доклинических моделях тройного отрицательного рак молочной железы. Раки (Базель). 12: 71 . Просмотр статьи: Google Scholar |
62 | До К., Вильскер Д., Джи Дж., Злотт Дж., Пресноводный Т, Киндерс Р.Дж., Коллинз Дж., Чен А.П., Дорошоу Дж. Х. и Куммар С.: Фаза Я изучаю одноэлементный AZD1775 (MK-1775), ингибитор киназы Wee1, у пациентов с рефрактерными солидными опухолями.J Clin Oncol. 33: 3409–3415. 2015. Просмотр статьи: Google Scholar: PubMed / NCBI |
63 | До КТ, Хилл С.Дж., Кочупураккал Б., Супко Ю.Г., Гэннон К., Андерсон А., Музикански А., Волански А., Хедглин Дж., Пармар K, et al: Abstract CT232: Фаза I комбинированного исследования CHK1 ингибитор прексасертиб (LY2606368) и олапариб у пациентов с серозный рак яичников высокой степени злокачественности и другие запущенные солидные опухоли. В: Proceedings of the American Association for Cancer Research. Ежегодное собрание 2019.Cancer Res. 79 (Приложение 13): Реферат № CT232. 2019. |
64 | Пэн В., Чен Дж.К., Лю С., Малу С., Кризи К., Tetzlaff MT, Xu C, McKenzie JA, Zhang C, Liang X и др.: Потеря PTEN способствует устойчивости к Т-клеточной иммунотерапии. Рак Discov. 6: 202–216. 2016. Просмотр статьи: Google Scholar: |
65 | Андре Ф, Сируелос Э, Рубовски Г, Кампоне М., Лойбл С., Руго Х.С., Ивата Х., Конте П., Майер И.А., Кауфман Б. и др.: Алпелисиб для PIK3CA-мутантного гормонального рецептора продвинутого уровня рак молочной железы.N Engl J Med. 380: 1929–1940. 2019. Просмотр статьи: Google Scholar |
66 | Ким С.Б., Дент Р., Им С.А., Эспье М., Блау С., Тан А.Р., Исакофф С.Дж., Оливейра М., Саура С., Вонгченко М.Дж. и др.: Ипатасертиб плюс паклитаксел по сравнению с плацебо плюс паклитаксел как терапия первой линии при метастатическом тройном отрицательном раке молочной железы (LOTUS): многоцентровый, рандомизированный, двойной слепой, плацебо-контролируемое исследование фазы 2. Ланцет Онкол. 18: 1360–1372. 2017. Просмотр статьи: Google Scholar: PubMed / NCBI |
67 | Dent R, Im SA, Espie M, Blau S, Tan AR, Исакофф С.Дж., Оливейра М., Саура С., Вонгченко М., Капп А.В. и другие: Обновление общей выживаемости (ОС) двойного слепого плацебо (PBO) контролируемое рандомизированное исследование LOTUS фазы 2 первой линии ипатасертиб (IPAT) + паклитаксел (PAC) для местного применения распространенный / метастатический тройной отрицательный рак молочной железы (mTNBC).J Clin Онкол. 36: 10082018. Просмотр статьи: Google Scholar |
68 | Дент Р., Ким С.Б., Оливейра М., Исакофф С.Дж., Барриос СН, О’Шонесси Дж., Лу Икс, Вонгченко М, Брэдли Д., Мани А, и др.: IPATunity130: основное рандомизированное исследование III фазы. оценка ипатасертиб (IPAT) + паклитаксел (PAC) для PIK3CA / AKT1 / PTEN-измененный продвинутый трижды отрицательный (TN) или гормональный рецептор-положительный HER2-отрицательный (HR + / HER2-) рак молочной железы (BC). J Clin Oncol. 36 (Дополнение 15): TPS11172018.Просмотр статьи: Google Scholar |
69 | Шмид П., Абрахам Дж., Чан С., Уитли Д., Брант А.М., Немсадзе Г., Бэрд Р.Д., Парк Ю.Х., Холл П.С., Перрен Т. и др.: Капивасертиб плюс паклитаксел по сравнению с плацебо плюс паклитаксел как терапия первой линии при метастатическом тройном отрицательном раке молочной железы: Испытание ПАКТ. J Clin Oncol. 38: 423–433. 2020. Просмотр статьи: Google Scholar |
70 | Шмид П., Кортес Дж., Робсон М., Ивата Х., Хегг Р., Верма С., Нечаева М., Сю Б., Хаддад В., Имедио Р. Э. и др.: Реферат OT2-08-02: Капивасертиб и паклитаксел в первой линии лечение пациентов с метастатическим тройным отрицательным молоком рак: исследование III фазы (CAPItello-290).В: Труды Симпозиум по раку груди в Сан-Антонио, 2019 г. Cancer Res. 80 (Дополнение 4): Реферат № ОТ2-08-02. 2020. |
71 | Шмид П., Лойрат Д., Савас П., Эспиноза Э, Бони В., Итальяно А., Уайт С., Сингел М.С., Витана Н., Мани А. и др.: Исследование фазы Ib по оценке тройной комбинации ипатасертиба (IPAT), атезоли-зумаб (атезо) и паклитаксел (PAC) или nab-PAC как терапия первой линии (1 л) для местнораспространенных / метастатических тройной отрицательный рак груди (TNBC).В: Труды Ежегодное собрание Американской ассоциации исследований рака, 2019 г. Cancer Res. 79 (Приложение 13): Реферат № CT049. 2019. |
72 | Ханахан Д. и Вайнберг Р.А.: Признаки рак: следующее поколение. Клетка. 144: 646–674. 2011. Просмотр статьи: Google Scholar: PubMed / NCBI |
73 | Дин Дж. Л., МакКлендон А. К. и Кнудсен Е. С.: Модификация ответа на повреждение ДНК терапевтическим CDK4 / 6 торможение.J Biol Chem. 287: 29075–29087. 2012. Просмотр статьи: Google Scholar: PubMed / NCBI |
74 | Cretella D, Fumarola C, Bonelli M, Alfieri R, La Monica S, Digiacomo G, Cavazzoni A, Galetti M, Generali D и Петронини PG: предварительное лечение ингибитором CDK4 / 6 пальбоциклибом улучшает эффективность паклитаксела в клетках TNBC. Научный представитель 9: 130142019. Просмотр статьи: Google Scholar: PubMed / NCBI |
75 | Кларк А.С., МакЭндрю Н.П., Троксель А., Фельдман М., Лал П., Розен М., Баррелл Дж., Редлингер К., Галлахер М., Брэдбери AR, et al: Комбинация паклитаксела и палбоциклиба: результаты испытание фазы I при запущенном раке груди.Clin Cancer Res. 25: 2072–2079. 2019. Просмотр статьи: Google Scholar: PubMed / NCBI |
76 | Тан А.Р., Райт Г.С., Туммала А.Р., Дансо М.А., Попович Л., Плуард Т.Дж., Хан Х.С., Войнович Ž, Васев Н., Ма Л. и др.: Трилациклиб плюс химиотерапия по сравнению с одной химиотерапией у пациентов с метастатическим тройным отрицательным раком груди: многоцентровое, рандомизированное открытое исследование фазы 2. Ланцет Онкол. 20: 1587–1601. 2019. Просмотр статьи: Google Scholar: PubMed / NCBI |
77 | Стайн З.Э., Уолтон З.е., Альтман Б.Дж., Ше А.Л. и Dang CV: MYC, метаболизм и рак.Рак Discov. 5: 1024–1039. 2015. Просмотр статьи: Google Scholar: PubMed / NCBI |
78 | Амбросио С, Аменте С, Наполитано Джи, Ди Palo G, Lania L и Majello B: MYC ухудшает разрешение сайт-специфическая репарация двухцепочечных разрывов ДНК. Mutat Res. 774: 6–13. 2015. Просмотр статьи: Google Scholar: PubMed / NCBI |
79 | Wiegmans AP, Al-Ejeh F, Chee N, Yap PY, Горски Дж. Дж., Да Силва Л., Болдерсон Е., Ченевикс-Тренч Дж., Андерсон Р., Simpson PT и др.: Rad51 поддерживает тройной отрицательный рак груди метастаз.Oncotarget. 5: 3261–3272. 2014. Просмотреть статью: Google Scholar: PubMed / NCBI |
80 | Кэри JPW, Каракас К., Буй Т, Чен Икс, Виджаярагхаван С., Чжао Ю., Ван Дж., Микуле К., Литтон Дж. К., Хант К. К. и Keyomarsi K: синтетическая летальность ингибиторов PARP в комбинации с блокадой MYC не зависит от статуса BRCA при тройном отрицательном результате рак молочной железы. Cancer Res. 78: 742–757. 2018. Просмотр статьи: Google Scholar: |
81 | Хориучи Д., Кусдра Л., Хаски СВ, Чандриани С., Ленбург М.Э., Гонсалес-Ангуло А.М., Кризман К.Дж., Базаров AV, Smyth JW, Davis SE и др.: Активация пути MYC в трижды отрицательный рак молочной железы является синтетическим летальным исходом с CDK торможение.J Exp Med. 209: 679–696. 2012. Просмотр статьи: Google Scholar: PubMed / NCBI |
82 | Hossain DMS, Javaid S, Cai M, Zhang C, Савант А., Хинтон М., Сате М., Грейн Дж., Блюменшейн В., Пинейро Э.М. и Chackerian A: динациклиб вызывает гибель иммуногенных клеток и усиливает подавление опухоли, опосредованное антителами к PD1. J Clin Invest. 128: 644–654. 2018. Просмотр статьи: Google Scholar: PubMed / NCBI |
83 | Чиен А.Дж., Глива А.С., Рахмапутри С., Диттрих HF, Majure MC, Rugo HS, Melisko ME, Munster PN, Park JW, Moasser MM, et al: фаза Ib испытания циклин-зависимой киназы ингибитор динаци-клиб (дина) в комбинации с пембролизумабом (П) у пациентов с запущенным тройным отрицательным раком молочной железы (TNBC) и корреляция ответа с гиперэкспрессией MYC.J Clin Oncol. 38 (1076) 2020. Просмотр статьи: Google Scholar |
84 | Коно М., Фудзи Т., Лим Б., Карутури М.С., Трипатия D и Уэно NT: функция рецепторов андрогенов и андрогены рецепторно-таргетная терапия при раке груди: обзор. JAMA Oncol. 3: 1266–1273. 2017. Просмотр статьи: Google Scholar: PubMed / NCBI |
85 | Герратана Л., Базиль Д., Буоно Дж., Де Пласидо S, Джулиано М, Миничилло С, Коину А, Марторана Ф, Де Санто I, Дель Мастро Л. и др.: Рецепторы андрогенов в тройной отрицательной груди рак: потенциальная цель для бесцельного подтипа.Лечение рака Откр. 68: 102–110. 2018. Просмотр статьи: Google Scholar: PubMed / NCBI |
86 | Anestis A, Zoi I, Papavassiliou AG и Карамузис М.В.: Рецепторы андрогенов при раке молочной железы — клинические и выводы доклинических исследований. Молекулы. 25 (358) 2020. Просмотр статьи: Google Scholar: PubMed / NCBI |
87 | Масуда Х., Баггерли К.А., Ван И, Чжан И, Гонсалес-Ангуло А.М., Мерик-Бернстам Ф., Валеро В., Леманн Б.Д., Пиетенпол Дж. А., Хортобадьи Г. Н. и др.: Дифференциальный ответ на неоадъювантная химиотерапия среди 7 случаев тройного отрицательного рака груди молекулярные подтипы.Clin Cancer Res. 19: 5533–5540. 2013. Просмотр статьи: Google Scholar: PubMed / NCBI |
88 | Thike AA, Yong-Zheng Chong L, Cheok PY, Li HH, Wai-Cheong Yip G, Huat Bay B, Tse GM, Iqbal J и Tan PH: Потеря экспрессии андрогенных рецепторов предсказывает раннее рецидивирование тройной отрицательный и базальный рак груди. Мод Pathol. 27: 352–360. 2014. Просмотр статьи: Google Scholar |
89 | Эчаваррия I, Лопес-Тарруэлла С, Пикорнелл A, Гарсиа-Саенс Х.А., Херес Й., Хоадли К., Гомес Х.Л., Морено Ф., Доктор медицины Монте-Миллан, Маркес-Родас I и др.: Патологический ответ в когорта с тройным отрицательным раком молочной железы, получавшая неоадъювант карбоплатин и доцетаксел в соответствии с усовершенствованным методом Леманна. классификация.Clin Cancer Res. 24: 1845–1852. 2018. Просмотр статьи: Google Scholar: PubMed / NCBI |
90 | Сантоха А, Санчес-Муньос А, Луч А, Chica-Parrado MR, Albanell J, Chacón JI, Antolín S, Jerez JM, de la Хаба Дж., Де Люк В. и др.: Подтипы тройного отрицательного рака груди. и частота патологического полного ответа на неоадъювантную химиотерапию. Oncotarget. 9: 26406–26416. 2018. Просмотр статьи: Google Scholar: PubMed / NCBI |
91 | Венема К.М., Бенсе Р.Д., Стинбрюгген Т.Г., Nienhuis HH, Qiu SQ, van Kruchten M, Brown M, Tamimi RM, Hospers GAP, Schröder CP и др.: Рассмотрение подтипа рака груди в нацеленный на рецептор андрогенов.Pharmacol Ther. 200: 135–147. 2019. Просмотр статьи: Google Scholar: PubMed / NCBI |
92 | Райс М.А., Малхотра С.В., Стоянова Т: Антиандрогены второго поколения: от открытия к стандарту лечения при кастрационно-резистентном раке простаты. Фасад Онкол. 9: 8012019. Просмотр статьи: Google Scholar: PubMed / NCBI |
93 | Гукальп А., Толани С., Исакофф С.Дж., Ингл Дж. Н., Лю М.К., Кэри Л.А., Блэквелл К., Руго Х., Набелл Л., Фореро А. и др.: Фаза II исследования бикалутамида у пациентов с андрогенами рецептор-положительный, эстроген-отрицательный метастатический рак груди рак.Clin Cancer Res. 19: 5505–5512. 2013. Просмотр статьи: Google Scholar: PubMed / NCBI |
94 | Гукальп А., Эдельвейс М, Патил С., Гундер М.М., Фейгин К.Н., Корбен А., Арумов А. и Трайна Т.А.: Реферат P3-11-04: Фаза I / II исследования палбоциклиба в комбинации с бикалутамидом для лечения андрогенных рецепторов (АР) + метастатическая грудь рак (МБК). В: Труды рака груди в Сан-Антонио, 2017 г. Симпозиум. Cancer Res 2018. 78 (Приложение 4): Аннотация № P3-11-04.2018. |
95 | Гукальп А, Бойл Л.А., Алано Т, Арумов А, Гаундер М.М., Патил С., Фейгин К., Эдельвейс М., Д’Андреа Г., Бромберг Дж., и др.: Фаза II испытания бикалутамида в комбинации с палбоциклиб для лечения метастазов по рецепторам андрогенов (+) рак молочной железы. J Clin Oncol. 38: 2020. Просмотр статьи: Google Scholar |
96 | Боннефой Х, Греллети Т, Тредан О, Сагатчян М., Даленс Ф., Майлез А., Л’Харидон Т., Котту П., Abadie-Lacourtoisie S, You B и др.: Исследование II фазы абиратерона ацетат плюс преднизон у пациентов с трижды отрицательный рецептор андрогенов положительный местнораспространенный или метастатический рак молочной железы (UCBG 12-1).Энн Онкол. 27: 812–818. 2016 г. Просмотр статьи: Google Scholar: PubMed / NCBI |
97 | Гукальп А., Дансо М.А., Элиас А.Д., Бардиа А., Али Х.Й., Поттер Д., Габрейл Нью-Йорк, Хейли Б.Б., Хонг Х.Т., Райли Е.К. и др.: Фаза (Ph) 2 стадия 1 клиническая активность севитеронела, селективного Ингибитор CYP17-лиазы и рецепторов андрогенов (AR) у женщин с продвинутый AR + тройной отрицательный рак груди (TNBC) или эстроген рецептор (ER) + BC: CLARITY-01. J Clin Oncol. 35: 11022017.Просмотр статьи: Google Scholar |
98 | Бардия А, Гукалп А, ДаКоста Н, Габраил Н, Дансо М., Али Х., Блэквелл К.Л., Кэри Л.А., Эйснер-младший, Баскин-Бей Е.С. и Traina TA: Фаза 1 исследования севитеронела, селективного CYP17 ингибитор рецепторов лиазы и андрогенов у женщин с эстрогенами рецептор-положительный или тройной отрицательный рак груди. Рак молочной железы Res Treat. 171: 111–120. 2018. Просмотр статьи: Google Scholar: PubMed / NCBI |
99 | Трэйна Т.А., Миллер К., Ярдли Д.А., Икл Дж., Шварцберг Л.С., О’Шонесси Дж., Градишар В., Шмид П., Винер Э., Келли С. и др.: Энзалутамид для лечения андрогенов. рецептор-экспрессирующий тройной отрицательный рак молочной железы.J Clin Oncol. 36: 884–890. 2018. Просмотр статьи: Google Scholar: PubMed / NCBI |
100 | Дент Р., Шмид П., Кортес Дж., Ким С.Б., Андре Ф., Абрамсон В., Кардосо Ф., Коллеони М., Моррис П., Стейнберг Дж. И др.: Реферат OT3-02-02: ENDEAR: рандомизированная международная фаза 3 исследование, сравнивающее эффективность и безопасность энзалутамида при комбинация с химиотерапией паклитакселом или в качестве монотерапии по сравнению с плацебо с паклитакселом у пациентов с диагностически-положительный тройной отрицательный рак груди.Cancer Res. 77: Реферат ОТ3-02-02. 2017. |
101 | Леманн Б.Д., Абрамсон В.Г., Сандерс М.Э., Майер ЭЛ, Хаддад ТК, Нанда Р., Ван Познак К., Сторниоло А.М., Нангиа-младший, Гонсалес-Эрикссон PI и др.: Многоцентровое исследование TBCRC 032 IB / II: Молекулярное понимание эффективности антагонистов AR и ингибиторов PI3K в пациенты с AR + метастатической тройной отрицательной молочной железы рак. Clin Cancer Res. 26: 2111–2123. 2020. Просмотр статьи: Google Scholar |
102 | Гилевски Т., Рагупати Дж., Бхута С., Уильямс LJ, Musselli C, Zhang XF, Bornmann WG, Спасова M, Bencsath KP, Панагеас К.С. и др.: Иммунизация метастатического рака груди. пациенты с полностью синтетическим конъюгатом globo H: испытание фазы I.Proc Natl Acad Sci USA. 98: 3270–3275. 2001. Просмотр статьи: Google Scholar: PubMed / NCBI |
103 | Хуан К.С., Юй А.Л., Цзэн Л.М., Чоу LWC, Хоу MF, Hurvitz SA, Schwab RB, Wong CH, Murray JL, Chang SC и др .: Рандомизированное исследование фазы II / III активной иммунотерапии с OPT-822 / OPT-821 у пациентов с метастатическим раком молочной железы. J Clin Онкол. 34 (Дополнение 15): S10032016. Просмотр статьи: Google Scholar |
104 | Бардия А, Майер И.А., Вахдат Л.Т., Толаней С.М., Исакофф С.Дж., Даймонд-младший, О’Шонесси Дж., Моруз Р.Л., Сантин А.Д., Абрамсон В.Г. и др.: Сацитузумаб говитекан-хзий в огнеупорных материалах. метастатический тройной отрицательный рак груди.N Eng J Med. 380: 741–751. 2019. Просмотр статьи: Google Scholar |
105 | Моди С, Пуштай Л., Фореро А, Мита М, Миллер К.Д., Вайз А., Беррис Х. III, Калински К., Цай М., Лю М.К. и др. al: Abstract PD3-14: Фаза 1 исследования конъюгата антитело-лекарственное средство SGN-LIV1A у пациентов с тройным отрицательным метастатический рак груди. Cancer Res. 78: 2018. |
106 | Han HS, Alemany CA, Brown-Glaberman UA, Плард Т.Дж., Синха Р., Стерренберг Д., Албайн К.С., Башо Р.К., Биггс Д., Boni V и др.: SGNLVA-002: открытое исследование фазы Ib / II в одной группе. ладиратузумаба ведотина (LV) в сочетании с пембролизумабом для лечение первой линии пациентов с неоперабельными местнораспространенными или метастатический тройной отрицательный рак груди.J Clin Oncol. 37 (Доп. 15): TPS11102019. Просмотр статьи: Google Scholar |
107 | Моди С., Парк Х, Мурти Р.К., Ивата Х, Тамура К., Цурутани Дж., Морено-Аспития А., Дои Т., Сагара Й., Редферн С. и др. al: Противоопухолевое действие и безопасность трастузумаба Дерустекана в пациенты с распространенным раком молочной железы с низкой экспрессией HER2: результаты из исследования фазы Ib. J Clin Oncol. 38: 1887–1896. 2020. Просмотр статьи: Google Scholar: PubMed / NCBI |
108 | Моди С., Отани С., Ли СС, Ван К., Саксена К. и Кэмерон Д.А.: фаза III, многоцентровая, рандомизированная, открытая испытание [фам-] трастузумаба дерукстекана (DS-8201a) по сравнению с Выбор исследователя при раке молочной железы с низким уровнем HER2.J Clin Oncol. 37 (Дополнение 15): TPS11022019. Просмотр статьи: Google Scholar |
109 | Молинье Джи, Гейер ФК, Магней Ф.А., Маккарти A, Кендрик Х., Натраджан Р., Маккей А., Григориадис А., Тутт А., Эшворт А. и др.: BRCA1 базальный рак молочной железы происходит из предшественники просветного эпителия, а не из базальных стволовых клеток. Клетка Стволовая клетка. 7: 403–417. 2010. Просмотр статьи: Google Scholar: PubMed / NCBI |
110 | Бернардо GM, Бебек G, Ginther CL, Сайзмор СТ, Лозада К.Л., Мидлер Д.Д., Андерсон Л.А., Годвин А.К., Абдул-Карим Ф.В., Slamon DJ и Keri RA: FOXA1 подавляет молекулярный фенотип клетки базального рака молочной железы.Онкоген. 32: 554–563. 2013. Просмотр статьи: Google Scholar |
111 | Су И, Субэди А, Блуштейн-Кимрон Н, Савова В., Кшистанек М., Ли Л., Марусык А., Табассум Д.П., Зак А., Flacker MJ, et al: Слияние соматических клеток выявляет обширные неоднородность в базальном раке груди. Cell Rep. 11: 1549–1563. 2015. Просмотр статьи: Google Scholar: PubMed / NCBI |
112 | Цао Р, Ван Л., Ван Х, Ся Л., Эрдджумент-Бромаж Х, Темпст П., Джонс Р.С. и Чжан И .: роль гистон h4 — метилирование лизина 27 при сайленсинге Polycomb-группы.Наука. 298: 1039–1043. 2002. Просмотр статьи: Google Scholar: PubMed / NCBI |
113 | Ямагиши М и Учимару К: нацеливание на EZh3 в терапии рака. Curr Opin Oncol. 29: 375–381. 2017. Просмотр статьи: Google Scholar: PubMed / NCBI |
114 | Ян СС, ЛаБафф А, Вэй Й, Не Л., Ся В., Huo L, Yamaguchi H, Hsu YH, Hsu JL, Liu D и др.: Фосфорилирование ЭЖ3 по Т416 с помощью CDK2 способствует злокачественности тройного отрицательный рак груди.Am J Transl Res. 7: 1009–1020. 2015.PubMed / NCBI |
115 | Не Л., Вэй И, Чжан Ф, Сюй Ю. Х, Чан ЛК, Xia W, Ke B, Zhu C, Deng R, Tang J, et al: CDK2-опосредованный сайт-специфическое фосфорилирование EZh3 управляет и поддерживает тройной отрицательный рак груди. Nat Commun. 10: 51142019. Просмотр статьи: Google Scholar: PubMed / NCBI |
116 | Ян Х, Филлипс Д.Л., Фергюсон А.Т., Нельсон WG, Герман Дж. И Дэвидсон NE: Синергетическая активация функциональных рецептор эстрогена (ER) -альфа с помощью ДНК-метилтрансферазы и гистона ингибирование деацетилазы при ER-альфа-отрицательном раке молочной железы клетки.Cancer Res. 61: 7025–7029. 2001.PubMed / NCBI |
117 | Шарма Д., Саксена Н.К., Дэвидсон Н.Е. и Вертино ПМ: Восстановление чувствительности к тамоксифену в эстрогенах рецепторно-отрицательные клетки рака молочной железы: реактивированные тамоксифен-связанные ER набирает отличительные комплексы корепрессоров. Cancer Res. 66: 6370–6378. 2006. Просмотр статьи: Google Scholar: PubMed / NCBI |
118 | Коннолли Р.М., Ли Х., Янковиц Р.С., Чжан З., Рудек М.А., Джетер С.К., Слейтер С.А., Пауэрс П., Вольф А.С., Феттинг Дж. Х. и др. al: Комбинированная эпигенетическая терапия при распространенном раке груди с 5-азацитидин и энтиностат: национальный рак II фазы Институт / Поддержка исследования рака.Clin Cancer Res. 23: 2691–2701. 2017. Просмотр статьи: Google Scholar: |
119 | Андерберг К., Ли Х., Фредрикссон Л., Андрэ Дж., Бетсхольц К., Ли Х, Эрикссон У. и Пьетрас К.: паракринная передача сигналов. за счет фактора роста тромбоцитов-CC способствует росту опухоли путем рекрутирование связанных с раком фибробластов. Cancer Res. 69: 369–378. 2009. Просмотр статьи: Google Scholar: PubMed / NCBI |
120 | Росвалл П., Боччи М., Барточек М., Ли Х., Кристиансен Г., Янссон С., Лен С., Сьелунд Дж., Рид С., Ларссон С. и др. al: Микроэкологический контроль подтипа рака груди через передачу сигналов паракринного тромбоцитарного фактора роста-CC.Нат Med. 24: 463–473. 2018. Просмотр статьи: Google Scholar: PubMed / NCBI |
121 | Park JJH, Hsu G, Siden EG, Thorlund K и Миллс Э.Дж.: Обзор прецизионной онкологической корзины и зонта испытания для врачей. CA Cancer J Clin. 70: 125–137. 2020. Просмотр статьи: Google Scholar: PubMed / NCBI |
Вирусы как модуляторы функций митохондрий
Митохондрии — это многофункциональные органеллы, выполняющие различные функции, включая производство и распределение энергии, апоптоз, индукцию иммунного ответа хозяина и вызывающие заболевания и старение.Опосредованные митохондриями иммунные ответы могут быть эволюционной адаптацией, с помощью которой митохондрии могли предотвращать проникновение вторгающихся микроорганизмов, тем самым делая их неотъемлемой частью клетки. Это делает их мишенью для всех вторгающихся патогенов, включая вирусы. Вирусы либо индуцируют, либо ингибируют различные митохондриальные процессы высокоспецифическим образом, чтобы они могли реплицироваться и производить потомство. Некоторые вирусы кодируют гомологи Bcl2 для противодействия проапоптотическим функциям клеточных и митохондриальных белков.Другие модулируют поры перехода проницаемости и либо предотвращают, либо индуцируют высвобождение апоптотических белков из митохондрий. Вирусы, такие как вирус простого герпеса 1, истощают митохондриальную ДНК хозяина, а некоторые, такие как вирус иммунодефицита человека, захватывают митохондриальные белки хозяина, чтобы полностью функционировать внутри клетки-хозяина. Во всех этих процессах участвуют клеточные белки, митохондриальные белки и вирусспецифические белки. В этом обзоре будут обобщены стратегии, используемые вирусами для использования клеточных митохондрий для успешного размножения и производства потомства вируса.
1. Введение
1.1. Митохондрии
Митохондрии — это клеточные органеллы, обнаруженные в цитоплазме почти всех эукариотических клеток. Одна из их важных функций — производить и обеспечивать клетку энергией в виде АТФ, который помогает в правильном поддержании клеточных процессов, что делает их незаменимыми для клетки. Помимо того, что они действуют как источник энергии для клетки, они действуют как общая платформа для выполнения множества клеточных функций в нормальных клетках или клетках, инфицированных микроорганизмами.Митохондрии участвуют в старении [1, 2], апоптозе [3-7], регуляции клеточного метаболизма [4, 8], контроле клеточного цикла [9-11], развитии клетки [12-14], противовирусные реакции [15], передача сигнала [16] и заболевания [17–20].
Хотя все митохондрии имеют одинаковую архитектуру, они сильно различаются по форме и размеру. Митохондрии состоят из внешней митохондриальной мембраны, внутренней митохондриальной мембраны, межмембранного пространства (пространства между внешней и внутренней мембранами) и матрикса (пространства внутри внутренней митохондриальной мембраны).Наружная мембрана представляет собой гладкий фосфолипидный бислой, в который встроены различные типы белков [21]. Наиболее важными из них являются порины, которые позволяют свободно переносить (экспортировать и импортировать) молекулы (белки, ионы, питательные вещества и АТФ) менее 10 кДа через мембраны. Наружная мембрана окружает внутреннюю мембрану, создавая межмембранное пространство, которое содержит такие молекулы, как Cyt-C, SMAC / Diablo и эндонуклеаза G. Она также действует как буферная зона между внешней мембраной и внутренней мембраной митохондрий.Внутренняя мембрана сильно извита в структуры, называемые кристами, которые увеличивают площадь поверхности мембраны и являются местами респираторных комплексов. Внутренняя мембрана митохондрий обеспечивает свободный перенос кислорода и углекислого газа. Предполагается, что движение воды через мембраны контролируется белком канала аквапоринов [22, 23], хотя в отчете предполагается иное [24]. Матрица содержит ферменты для аэробного дыхания, растворенный кислород, воду, углекислый газ и перерабатываемые промежуточные продукты, которые служат в качестве энергетических челноков и выполняют другие функции.
Митохондрии содержат единственный кольцевой ДНК-геном размером 16 т.п.н., который кодирует 13 белков (в основном субъединицы дыхательных цепей I, II, IV и V), 22 митохондриальных тРНК и 2 рРНК [25, 26]. Митохондриальный геном не имеет оболочки (как ядерная оболочка), содержит мало интронов и не следует универсальному генетическому коду [27]. Хотя большинство митохондриальных белков кодируется ядерной ДНК и импортируется в митохондрии (обзор [21, 28–31]), митохондрии синтезируют мало белков, которые необходимы для их дыхательной функции [1, 27].
Белки, предназначенные для митохондрий, имеют либо внутренне локализованные [28], либо локализованные на аминоконцевых концах [21] пре-последовательности, известные как сигналы митохондрий / матриксной локализации (MLS), которые могут иметь длину от 10 до 80 аминокислот с преимущественно положительно заряженными аминокислотами. Комбинация этих предпоследовательностей с соседними участками определяет локализацию белка в соответствующих компартментах митохондрий. Наружная митохондриальная мембрана содержит два основных транслокатора, а именно: (а) транслоказу внешней мембраны (TOM) 40, которая функционирует как входные ворота для большинства митохондриальных белков с MLS, и (b) механизм сортировки и сборки (SAM) или транслоказа β -баррель (TOB) белок, который представляет собой специализированный аппарат для встраивания мембранных белков бета-ствола [32].Как только белки проходят через внешнюю мембрану, они рекрутируются с помощью транслоказо-ассоциированного мотора (PAM) в транслоказу комплексов внутренней митохондриальной мембраны (TIM) 23, которая опосредует импорт белков в матрицу. Наконец, пре-последовательности расщепляются в матриксе, а белки модифицируются до своей третичной структуры и становятся функциональными [30].
1.2. Вирусы
Вирусы — это бесклеточные облигатные внутриклеточные микроорганизмы, которые инфицируют живые клетки / организмы и являются единственным исключением из клеточной теории, предложенной Шлейденом и Шванном в 1838/1839 [33].Вирусы имеют внешний белковый капсид и ядро нуклеиновой кислоты. Обычно вирусные нуклеиновые кислоты могут быть либо ДНК (двух- или одноцепочечной), либо РНК (+ или — смысловой одноцепочечной или двухцепочечной РНК). Некоторые вирусы покрыты оболочкой из гликопротеинов. Вирусы издавна ассоциировались с живыми организмами, и именно в конце века было подробно изучено их взаимоотношение с различными клеточными органеллами. Чтобы выжить и размножаться в клетке, вирусам необходимо взять под контроль различные клеточные органеллы, участвующие в защитных и иммунных процессах.Им также требуется энергия для репликации и выхода из клетки. Попав внутрь клетки-хозяина, они модулируют различные клеточные сигнальные пути и органеллы, включая митохондрии, и используют их для собственного выживания и репликации. В этом обзоре обобщены функции митохондрий и то, как вирусы их модулируют (рис. 1).
2. Вирусы регулируют гомеостаз Ca
2+ в клетках-хозяевах2.1. Ca
2+ ГомеостазCa 2+ является одним из наиболее распространенных и универсальных элементов в клетке и действует как вторичный посредник, регулирующий многие клеточные процессы [34].Ранее считалось, что внешняя мембрана митохондрий проницаема для Ca 2+ , но недавние исследования показывают, что внешняя мембрана содержит потенциал-зависимые анионные каналы (VDAC), имеющие связывающие домены Ca 2+ , которые регулируют проникновение Ca 2+ в межмембранное пространство митохондрий [35–37]. Поступление Ca 2+ через внутреннюю мембрану регулируется митохондриальным унипортером Ca 2+ (MCU), который является высокоселективным каналом Ca 2+ , который регулирует поглощение Ca 2+ на основе митохондриальных мембранный потенциал (ММП).Чистое движение заряда из-за поглощения Ca 2+ прямо пропорционально уменьшению MMP [38]. Второй механизм, который помогает в перемещении Ca 2+ через мембрану митохондрий, называется механизмом захвата «быстрого режима» (RaM) [39]. В этом процессе Ca 2+ переносится через митохондриальную мембрану путем обмена с Na + , который, в свою очередь, зависит от его обмена с ионом H + и, следовательно, с ММП. Этот ионный обмен через митохондриальную мембрану снижает ММП, и его поддержание зависит от цепи переноса электронов (ETC).Третий механизм включает IP 3 R, канал Ca 2+ в эндоплазматическом ретикулуме. IP 3 R связан с митохондриальным VDAC через белок 75, регулирующий глюкозу (GRP75). Этот переход регулирует / облегчает обмен Ca 2+ от IP 3 R к VDAC [40].
Ca 2+ механизм оттока регулируется переходной порами проницаемости (PTP). PTP собирается во внутренней и внешней мембранах митохондрий [41, 42], с сайтами связывания Ca 2+ на матричной стороне внутренней мембраны.PTP регулирует высвобождение митохондриального Ca 2+ посредством строго регулируемого механизма «мерцания», который контролирует открытие и закрытие поры [43]. RaM работает синхронно с изоформой 1 рианодинового рецептора (RyR), которая является еще одним очень важным каналом высвобождения кальция [44]. И RyR, и RaM регулируют феномен связи возбуждения и метаболизма, при котором сокращение, индуцированное цитозольным Ca 2+ , сочетается с митохондриальной стимуляцией окс-фосфом Ca 2+ [45]. Однако перегрузка митохондрий Ca 2+ может привести к длительному открытию поры, что приводит к патологии [46].Хотя Ca 2+ участвует в активации многих клеточных процессов, включая стимуляцию АТФ-синтазы [47, 48], аллостерическую активацию ферментов цикла Кребса [49, 50] и транслоказы аденин-нуклеотидов (ANT) [51] , основная роль митохондриального Ca 2+ заключается в стимуляции окс-фосфора [52–54]. Таким образом, повышенное содержание Ca 2+ в митохондриях приводит к усилению регуляции всего механизма ох-фос, что затем приводит к более быстрой активности дыхательной цепи и более высокому выходу АТФ, что затем может удовлетворить потребность клеток в АТФ.Ca 2+ также активирует другие митохондриальные функции, включая активацию N-ацетилглутамин синтетазы с образованием N-ацетилглутамина [55], мощную аллостерическую активацию карбамоилфосфатсинтетазы и цикл мочевины [56]. Таким образом, любое нарушение митохондриального или цитозольного гомеостаза Ca 2+ имеет серьезные последствия для функции клеток. Более того, митохондриальный Ca 2+ , особенно в высоких концентрациях, наблюдаемых при патологии, по-видимому, оказывает несколько отрицательных эффектов на функции митохондрий [57].
2.2. Регуляция вирусами
Ряд вирусов изменяют регуляторную активность клетки Ca 2+ для их выживания. Вирус простого герпеса (HSV) 1 вызывает постепенное снижение (65%) поглощения митохондриальным Ca 2+ в течение 12 часов литического цикла [58], что способствует репликации вируса. Хотя митохондриальное поглощение Ca 2+ продолжает колебаться на протяжении всего периода инфицирования клеток вирусом кори, общее количество клеточного Ca 2+ остается неизменным [58], что свидетельствует о жестком контроле, который вирус оказывает на клеточные процессы во время его жизненный цикл.
Основной белок вируса гепатита С (HCV) нацелен на митохондрии и увеличивает Ca 2+ [59, 60]. Белок NS5A HCV вызывает изменения в гомеостазе Ca 2+ [61–63]. Оба эти белка могут быть ответственны за патогенез заболеваний печени, связанных с инфекцией ВГС. Даже в клетках, коинфицированных HCV и вирусом иммунодефицита человека (ВИЧ), эти вирусы усиливают активность MCU, вызывая клеточный стресс и апоптоз [59, 64]. Белок p7 HCV образует пориноподобные структуры [65] и вызывает приток Ca 2+ в цитоплазму из запасающих органелл [66].Эти белки HCV нарушают гомеостаз Ca 2+ на разных стадиях инфекции и, таким образом, помогают увеличить выживаемость клетки. Интересно, что взаимодействие протеина X вируса гепатита B (HBV) с VDAC вызывает высвобождение Ca 2+ из митохондрий / эндоплазматического ретикулума (ER) / golgi запасающих органелл в цитоплазматический компартмент, что, по-видимому, способствует репликации вируса [67, 68].
Белок Nef ВИЧ взаимодействует с IP 3 R [69] и индуцирует повышение цитозольного Ca 2+ за счет промотирования на независимую от Т-клеточного рецептора активацию пути NFAT [70].Активированный NFAT, в свою очередь, вызывает внутриклеточные колебания Ca 2+ с низкой амплитудой, способствуя транскрипции и репликации вирусного гена [71].
Ca 2+ является важным фактором для различных стадий жизненного цикла ротавируса и устойчивости к вириону ротавируса [72]. Белок NSP4 ротавируса увеличивает цитозольную концентрацию Ca 2+ за счет активации фосфолипазы C (PLC) и результирующего истощения ER Ca 2+ через IP 3 R [73, 74].Это изменение гомеостаза Ca 2+ было приписано увеличению проницаемости клеточной мембраны [75]. Сообщалось, что снижение клеточных концентраций Ca 2+ к концу жизненного цикла способствует высвобождению ротавируса из клетки [76].
Белок 2BC полиовируса увеличивает внутриклеточные концентрации Ca 2+ в клетках на 4 часа. После заражения, что необходимо для экспрессии вирусных генов [77, 78]. К концу жизненного цикла вируса высвобождение Ca 2+ из просвета ER через IP 3 R и RyR каналы вызывает накопление Ca 2+ в митохондриях через унипортер и VDAC, что приводит к дисфункции митохондрий и апоптоз [79].Напротив, белок 2B вируса Коксаки снижает проницаемость мембраны за счет снижения концентрации Ca 2+ в инфицированных клетках [80, 81] из-за его пориноподобной активности, которая приводит к оттоку Ca 2+ из органелл. Снижение трафика белка и низкая концентрация Ca 2+ в golgi и ER способствуют образованию комплексов репликации вируса, подавляют противовирусный иммунный ответ хозяина и подавляют апоптоз [82, 83].
Энтеровирусы организуют апоптотический процесс в течение своего жизненного цикла, чтобы улучшить его проникновение, выживание и высвобождение.Нарушение цитоплазматического гомеостаза Ca 2+ через 2–4 часа. постинфекция совпадает с подавлением апоптотического ответа, что можно объяснить снижением цитотоксических уровней Ca 2+ в клетке и митохондриях. Это также обеспечивает вирусу оптимальные условия для репликации и синтеза белка. Наконец, снижение уровней Ca 2+ в митохондриях и других запасающих органеллах (ER и golgi) вызывает повышение цитозольной концентрации Ca 2+ , что приводит к образованию везикул и гибели клеток, что способствует высвобождению вируса [81 , 84, 85].
Белок pUL37 × 1 цитомегаловируса человека (HCMV) локализуется в митохондриях [86] и вызывает перенос Ca 2+ из ER в митохондрии через 4-6 часов. После заражения [87]. Активное поглощение Ca 2+ митохондрией индуцирует продукцию АТФ и других зависимых от Ca 2+ ферментов, ускоряющих репликацию вируса, а снижение уровней Ca 2+ в ЭПР оказывает антиапоптотическое действие [88].
Белок 6,7K, кодируемый областью E3 HAdV-2, локализуется в ER и помогает поддерживать гомеостаз ER Ca 2+ в трансфицированных клетках, тем самым подавляя апоптоз [89].
3. Вирусы вызывают окислительный стресс в клетках-хозяевах
3.1. Электронная транспортная цепь
Дыхательная цепь митохондрий является основным и наиболее важным источником активных форм кислорода (RO) в клетке. Супероксид () — это первичная АФК, продуцируемая митохондриями. В нормальном состоянии утечка электронов между комплексами цепи переноса электронов (ETC) незначительна или отсутствует. Однако в стрессовых условиях небольшая часть электронов покидает комплекс III и достигает комплекса IV [90].Эта преждевременная утечка электронов к кислороду приводит к образованию двух типов супероксидов, а именно, в его анионной форме и в его протонированной форме.
Утечка электронов происходит в основном из сайтов Q O комплекса III, которые расположены непосредственно рядом с межмембранным пространством, что приводит к высвобождению супероксидов либо в матриксе, либо во внутреннем мембранном пространстве митохондрий [91–94]. Около 25–75% от общей утечки электронов через Комплекс III может составлять чистый внемитохондриальный выброс супероксида [95–97].Таким образом, основным источником в митохондриях является промежуточный радикал убисемихинона (QH • ), образующийся во время цикла Q в сайте Q O комплекса III [98–100]. Комплекс I также является источником АФК, но механизм генерации АФК менее ясен. Недавние сообщения предполагают, что глутатионилирование [101] или PKA-опосредованное фосфорилирование [101-103] комплекса I может повышать генерацию ROS. Обратный поток электронов от комплекса I к комплексу II также может приводить к образованию АФК [99].
Разнообразные клеточные защитные механизмы поддерживают постоянную концентрацию этих оксидантов на нетоксичных уровнях. Этот тонкий баланс между образованием АФК и метаболизмом может быть нарушен различными ксенобиотиками, включая вирусные белки. Основная причина образования АФК в инфицированных вирусом клетках — ограничение размножения вируса. Однако АФК также действует как сигнал для различных клеточных путей, и вирус использует хаос, созданный внутри клетки, для своей репликации.
3.2. Вирусы индуцируют активные формы кислорода
Ряд вирусов вызывают окислительный стресс в клетках-хозяевах, что прямо или косвенно помогает им выжить. Сообщалось, что аденовирус человека (HAdV-) 5 вызывает разрыв эндосомальной мембраны при инфекции, что приводит к высвобождению лизосомальных катепсинов, которые вызывают выработку АФК. Катепсины также вызывают разрушение митохондриальной мембраны, что приводит к высвобождению АФК из митохондрий, вызывая окислительный стресс [104].
Основной белок ВГС вызывает окислительный стресс в клетке и изменяет пути апоптоза [64, 105–107]. E1, E2, NS3 и коровый белок HCV являются мощными индукторами ROS и могут вызывать повреждение ДНК хозяина независимо [107, 108] или опосредовано оксидом азота (NO), таким образом способствуя репликации вируса.
АФК образуются при ВИЧ-инфекции [64, 109–111]. H 2 O 2 , АФК, генерируемые во время ВИЧ-инфекции, сильно индуцируют длинный концевой повтор ВИЧ (LTR) посредством активации NF-каппа B.Нарушение активности LTR отменяет активацию LTR в ответ на ROS, тем самым способствуя репликации вируса [112]. ВИЧ также вызывает обширное клеточное повреждение из-за увеличения выработки АФК и снижения выработки цитозольных антиоксидантов [113]. Коинфекция ВИЧ и HCV вызывает фиброз печени, прогрессирование которого регулируется посредством генерации ROS зависимым от NF- κ B образом [113].
Вирус Эпштейна-Барра (EBV) вызывает повышенный окислительный стресс в клетках-хозяевах в течение 48 часов.Во время литического цикла, указывающего на роль АФК в высвобождении вируса [114]. Окислительный стресс активирует ранний ген EBV BZLF-1, что вызывает реактивацию литического цикла EBV [114]. Было высказано предположение, что это играет важную роль в патогенезе EBV-ассоциированных заболеваний, включая злокачественные трансформации [115, 116].
Интересно, что HBV вызывает как увеличение, так и уменьшение окислительного стресса, что увеличивает его выживаемость в клетках-хозяевах [117, 118]. HBV вызывает сильную активацию регулируемых Nrf2 / ARE генов in vitro и in vivo посредством активации c-Raf и MEK белком X HBV, таким образом защищая клетки от индуцированного HBV окислительного стресса и способствуя возникновению инфекции [119 ].Белок X HBV также индуцирует опосредованную ROS активацию Forkhead box класса O4 (Foxo4), повышая устойчивость к гибели клеток, вызванной окислительным стрессом [120]. Однако отчеты также предполагают, что при воздействии окислительного стресса белок X HBV ускоряет потерю белка Mcl-1 через каскад каспазы-3, вызывая проапоптотические эффекты [118]. Коинфекция HCV также вызывает генотоксические эффекты в лимфоцитах периферической крови из-за повышенного окислительного повреждения и снижения MMP [121]. Возможно, что противоречивые функции протеина X холода HBV происходят на разных стадиях репликации вируса.
Вирус энцефаломиокардита (EMCV) вызывает окислительный стресс в клетках во время инфекции, повреждая нейроны, что является важным процессом в патогенезе инфекции EMCV [122].
4. Вирусы регулируют потенциал митохондриальной мембраны в клетках-хозяевах
4.1. Митохондриальный мембранный потенциал
Мембранный потенциал (МП) — это разница в напряжении или электрическом потенциале между внутренней и внешней стороной мембраны. Мембранный потенциал создается либо электрической силой (взаимное притяжение или отталкивание между положительными и отрицательными), либо диффузией частиц от высоких до низких концентраций.Потенциал митохондриальной мембраны (ММР) — это МП (180 мВ) через внутреннюю мембрану митохондрий, которая обеспечивает энергию для синтеза АТФ. Движение протонов от комплекса I к V электронной транспортной цепи (ETC), расположенной во внутренней митохондриальной мембране, создает электрический потенциал на внутренней мембране , , который важен для правильного поддержания производства ETC и ATP. Сообщаемые значения MMP для митохондрий ( in vivo ) различаются от вида к виду и от одного органа к другому в зависимости от функции митохондрий, белкового состава и степени активности окислительного фосфорилирования, необходимой в этой части тела [43].
Напряжение зависимые анионные каналы (VDACs), также известные как митохондриальные порины, образуют каналы во внешних митохондриальных мембранах и действуют как первичный путь для движения метаболитов через внешнюю мембрану [37, 96, 123–125]. Кроме того, ряд факторов, включая окислительный стресс, перегрузку кальцием и истощение АТФ, вызывают образование неспецифических пор перехода митохондриальной проницаемости (MPTP) во внутренней митохондриальной мембране, которая также отвечает за поддержание ММП [36, 37, 126 ].Внешняя мембрана VDAC, адениннуклеотидтранслоказа внутренней мембраны (ANT) [127] и циклофилин D (CyP-D) в матриксе являются структурными элементами поры перехода митохондриальной проницаемости (MPTP).
В открытом состоянии MPTP увеличивает проницаемость внутренней митохондриальной мембраны для ионов и растворенных веществ до 1,5 кДа, что вызывает диссипацию MMP и диффузию растворенных веществ вниз по градиентам их концентрации с помощью процесса, известного как переход проницаемости [128, 129 ]. Открытие МРТР сопровождается осмотическим потоком воды, пассивным набуханием, разрывом внешней мембраны и высвобождением проапоптотических факторов, ведущих к гибели клетки [42, 130].Из-за последующего истощения АТФ и дерегуляции Ca 2+ , открытие MPTP было предложено как ключевой элемент в определении судьбы клетки до того, как была предложена роль митохондрий в апоптозе [129].
MMP может быть изменен множеством стимулов, включая внезапный всплеск ROS [43, 107], перегрузку Ca 2+ в митохондриях или клетке [48, 57, 131] и / или белками вторжения. вирусы [109, 132, 133]. Как правило, увеличение или уменьшение ММП связано с индукцией или предотвращением апоптоза соответственно.Предотвращение апоптоза на ранних стадиях вирусной инфекции — обычная стратегия, используемая вирусами для предотвращения иммунного ответа хозяина и стимулирования их репликации. Напротив, индукция апоптоза на более поздних стадиях вирусной инфекции — это стратегия, используемая вирусами для высвобождения дочерних вирионов для распространения в окружающие клетки.
4.2. Регуляция вирусами
Многие вирусные белки изменяют проницаемость митохондриальных ионов и / или мембранный потенциал для их выживания в клетке.Р7, гидрофобная интегральная мембрана [134] виропротеин [135] ВГС, локализуется в митохондриях [66] и контролирует проницаемость мембраны для катионов [66, 136], способствуя выживанию клеток для репликации вируса [135].
Белок R (Vpr) ВИЧ, небольшой дополнительный белок, локализуется в митохондриях, взаимодействует с ANT, модулирует MPTP и вызывает потерю MMP, способствуя высвобождению Cyto C [137], что приводит к гибели клеток [138, 139] . Белок Tat ВИЧ также модулирует MPTP, что приводит к накоплению Tat в митохондриях и индукции потери MMP, что приводит к каспазозависимому апоптозу [140].
Белок M11L поксвируса миксомы локализуется в митохондриях, взаимодействует с митохондриальным периферическим бензодиазепиновым рецептором (PBR) и регулирует MPTP [141], ингибируя потерю MMP [142] и таким образом подавляя индукцию апоптоза во время вирусной инфекции [143]. Белок FIL вируса осповакцины подавляет регуляцию проапоптотического белка семейства Bcl-2 Bak и ингибирует потерю MMP и высвобождение Cyt-C [144, 145]. Белок crmA / Spi-2 вируса коровьей оспы, ингибитор каспазы 8, модулирует МРТР, предотвращая, таким образом, апоптоз [146].
Белок PB1-F2 вирусов гриппа A локализуется в митохондриях [147–150] и взаимодействует с VDAC1 и ANT3 [151], что приводит к снижению MMP, что вызывает высвобождение проапоптотических белков, вызывающих гибель клеток. Недавние данные показывают, что PB1-F2 также способен формировать поры неселективных белковых каналов, что приводит к изменению морфологии митохондрий, диссипации MMP и гибели клеток [150]. Белок M2 вируса гриппа, виропротеин, вызывает изменение морфологии митохондрий, диссипацию ММП и гибель клеток (обзор [135]).
Дополнительный белок p13II, кодируемый ORF x-II человеческого Т-лимфотропного вируса (HTLV), нового члена семейства виропротеинов [152], локализуется в митохондриях инфицированных клеток и увеличивает количество ММП, что приводит к апоптозу [ 153] и набухание митохондрий [153–155].
Белок Orf C вируса дермальной саркомы судака (WDSV) локализуется в митохондриях [156] и вызывает перинуклеарную кластеризацию митохондрий и потерю MMP [156], что приводит к высвобождению проапоптотических факторов, вызывая, таким образом, апоптоз.
Белок 2B вируса Коксаки снижает ММП за счет снижения концентрации Ca 2+ в инфицированных клетках [80, 81].
5. Вирусы регулируют апоптоз
5.1. Апоптоз
Во время совместной эволюции вирусов с их хозяевами вирусы разработали несколько стратегий манипулирования аппаратом клетки-хозяина для их выживания, репликации и высвобождения из клетки. Вирусы нацелены на клеточный аппарат апоптоза на критических стадиях репликации вируса, чтобы достичь своих целей [157, 158].В зависимости от необходимости вирус может ингибировать [159] или индуцировать [160] апоптоз с очевидной целью репликации и распространения соответственно [158, 159]. Нарушение функции митохондрий может вызвать гибель клеток из-за нарушения регуляции сигнальных путей Ca 2+ и истощение АТФ или апоптоз из-за регуляции белков семейства Bcl-2. Апоптоз — это запрограммированная гибель клеток [161], характеризующаяся образованием пузырей на мембранах, конденсацией ядра и цитоплазмы и расщеплением эндонуклеосомной ДНК.Процесс начинается, как только клетка ощущает физиологические или стрессовые стимулы, что нарушает гомеостаз клетки [162, 163]. Апоптотическая смерть клетки может рассматриваться как врожденная реакция на ограничение роста микроорганизмов, включая вирусы, атакующие клетку.
Два основных пути, а именно внешний и внутренний, участвуют в запуске апоптоза [163, 164]. Внешний путь опосредуется передачей сигналов через рецепторы смерти, такие как фактор некроза опухоли или рецептор Fas-лиганда, вызывая сборку индуцирующего смерть сигнального комплекса (DISC) с привлечением таких белков, как каспазы, что приводит к проницаемости митохондриальной мембраны.Во внутреннем пути сигналы действуют непосредственно на митохондрии, что приводит к пермеабилизации митохондриальной мембраны до того, как каспазы активируются, вызывая высвобождение Cyt-C [165, 166], который рекрутирует APAF1 [167, 168], что приводит к прямой активации каспазы 9 [ 35, 169]. И внешние, и внутренние процессы объединяются при активации нижестоящих эффекторных каспаз (т.е. каспазы-3) [170], которая ответственна за индукцию морфологических изменений, наблюдаемых в апоптотической клетке.Помимо Cyt-C, Smac / DIABLO, а также независимые от каспаз эффекторы, индуцирующие гибель, фактор (AIF) и эндонуклеаза G [171–173] действуют как активатор каспазы.
Семейство белков B-клеточной лимфомы (Bcl-) 2 жестко регулирует апоптотические процессы, вовлекающие митохондрии [174, 175]. К настоящему времени описано более 20 белков семейства Bcl-2 млекопитающих [176, 177]. Они были классифицированы по наличию доменов гомологии Bcl-2 (BH), расположенных в порядке Bh5-Bh4-Bh3-Bh2, и С-концевого гидрофобного трансмембранного (TM) домена, который прикрепляет их к внешней митохондриальной мембране [178] .Высококонсервативные домены Bh2 и Bh3 ответственны за антиапоптотическую активность и мультимеризацию белков семейства Bcl-2. Домен Bh4 в основном отвечает за проапоптотическую активность, а менее консервативный домен Bh5 необходим для антиапоптотической активности Bcl-2 и белков [174, 178]. Большинство антиапоптотических белков являются мультидоменными белками, которые содержат все четыре домена BH (от Bh2 до Bh5) и домен TM. Напротив, проапоптотические белки являются либо мультидоменными белками, которые содержат три домена BH (от Bh2 до Bh4), либо однодоменными белками, которые содержат один домен (Bh4) [158].Белки Bcl-2 регулируют MMP в зависимости от того, принадлежат ли они к про- или антиапоптотической ветви семейства, соответственно. ММП отмечает тупик апоптоза, за которым клеткам суждено умирать [125, 166, 179–183].
5.2. Регуляция вирусами
Вирусы кодируют гомологи белков Bcl-2 (vBcl-2), которые могут вызывать (проапоптотический) или предотвращать (антиапоптотический) апоптоз, тем самым помогая вирусам завершить свой жизненный цикл в клетках-хозяевах [117, 163, 175]. В то время как vBcl-2s и клеточные Bcl-2s обладают ограниченной гомологией последовательностей, их вторичные структуры, по прогнозам, будут весьма сходными [158, 174, 184].Во время первичной инфекции взаимодействие между vBcl-2 и другими белками увеличивает продолжительность жизни клеток-хозяев, что приводит к эффективному продуцированию вирусного потомства и, в конечном итоге, к распространению инфекции на новые клетки. Это также способствует сохранению вируса в клетках, позволяя латентно инфицирующим вирусам перейти в продуктивную инфекцию. Пути и стратегии, используемые вирусами для индукции / ингибирования апоптоза, были рассмотрены ранее [185].
Многие вирусы кодируют гомологи антиапоптотических белков Bcl-2, которые преимущественно локализуются в митохондриях и могут взаимодействовать с другими проапоптотическими гомологами Bax.E1B19K, кодируемый аденовирусом человека (HAdV-) 5, содержит Bh2 и Bh4-подобные домены и блокирует TNF-альфа-опосредованную передачу сигналов смерти путем ингибирования формы Bax, которая прерывает активацию каспазы ниже каспазы-8 и выше каспазы- 9 [186, 187]. Подобно HAdV-5 E1B19K [186], некоторые вирусы кодируют гомологи Bcl-2, лишенные домена Bh5, которые, как полагают, действуют путем ингибирования проапоптотических членов белков семейства Bcl-2. Белок FPV309, кодируемый вирусом оспы птиц, содержит высококонсервативные Bh2- и Bh3-подобные домены, а также скрытый домен Bh4, взаимодействует с белком Bax и ингибирует апоптоз [188].Белок A179L, кодируемый вирусом африканской чумы свиней (ASFV), содержит домены Bh2 и Bh3, взаимодействует с белками Bax-Bak и ингибирует апоптоз [189, 190]. Гомолог Bcl-2 (vBcl-2), кодируемый герпесвирусом saimiri (HVS), содержит Bh4 и Bh5-подобные домены и взаимодействует с Bax, таким образом стабилизируя митохондрии против различных апоптотических стимулов, предотвращая гибель клеток [191]. ORF E4, кодируемая лошадьми Herpesvirus-3, содержит домены Bh2 и Bh3 [192], которые могут взаимодействовать с Bax и иметь важное значение для антиапоптотической активности [193].
Вирусы также кодируют гомологи проапоптотических белков Bcl-2. HBV кодирует белок X, белок vBcl-2, содержащий Bh4, который локализуется в митохондриях и взаимодействует с VDAC, вызывая потерю MMP, что приводит к апоптозу [117, 121, 194, 195], или взаимодействует с Hsp60 и индуцирует апоптоз [196 ]. Напротив, другое исследование выявило защитные эффекты HB-X в ответ на проапоптотические стимулы (Fas, TNF и синдром отмены сыворотки), но не на химические апоптотические стимулы [197].Известно, что белок X HBV стимулирует NF κ B [198, 199], SAPK [200, 201] и PI3K / PKB [202] для предотвращения апоптоза. Возможно, что различные функции протеина X HBV происходят в разное время цикла репликации вируса в инфицированных клетках. Белок BALF1, кодируемый EBV, содержит домены Bh2 и Bh5 [203], которые взаимодействуют с белками Bax-Bak [192] и ингибируют антиапоптотическую активность BHRF1 EBV и белка Bcl-2 вируса саркомы Капоши (KSV), оба из которых являются которые содержат домены Bh2 и Bh3 [204] и взаимодействуют только с белками Bh4 [205].
Таким образом, эффекты вирусных гомологов Bcl-2, по-видимому, сосредоточены вокруг митохондрий и включают предотвращение или индукцию потери ММП. Индукция потери MMP приводит к высвобождению Cyto C и других проапоптотических сигналов в цитозоль и активации нижележащих каспаз, что приводит к гибели клеток и распространению вирусов в соседние клетки для дальнейшей инфекции.
Вирусы кодируют про / антиапоптотические белки, которые не проявляют гомологии с белками Bcl-2 [158]. Белок E6 вируса папилломы человека (HPV) подавляет сигнал Bax перед митохондриями [206, 207] и предотвращает высвобождение Cyto C, AIF и Omi, предотвращая, таким образом, апоптоз [208].Эта активность E6 по отношению к другому проапоптотическому белку семейства Bcl2, Bak, является ключевым фактором, способствующим выживанию HPV-инфицированных клеток, что, в свою очередь, способствует завершению жизненного цикла вируса [207]. Энтеровирус (EV) 71 индуцирует конформационные изменения Bax и увеличивает его экспрессию в клетках после инфекции и индуцирует активацию каспаз 3, 8 и PARP, вызывая каспазозависимый апоптоз [209]. Напротив, капсид вируса краснухи связывается с Bax, образует олигогетеромеры и предотвращает образование пор на митохондриальной мембране, тем самым предотвращая индуцированный Bax апоптоз [210].
Вирусы также кодируют белки, которые действуют как вирусные митохондриальные ингибиторы апоптоза (vMIA), таким образом защищая клетки. Вариант сплайсинга UL37 HCMV действует как vMIA и защищает клетки от апоптоза [211], тем самым помогая вирусам завершить цикл репликации. Он локализуется в митохондриях и взаимодействует с ANT [211] и Bax [212, 213]. VMIA HCMV имеет N-концевой домен митохондриальной локализации и C-концевой антиапоптотический домен [211], который рекрутирует Bax в митохондрии и предотвращает потерю MMP.Он защищает клетки от лигирования CD95 [211] и гибели клеток, вызванной окислительным стрессом [214, 215], и предотвращает слияние митохондрий [216], тем самым способствуя выживанию клеток.
vMIA не ингибирует апоптотические события перед митохондриями, но может влиять на такие события, как сохранение генерации АТФ, ингибирование высвобождения Cyto C и активация каспазы 9 после индукции апоптоза. Однако точные механизмы событий вокруг vMIA все еще остаются под вопросом.
6.Вирусы модулируют митохондриальный противовирусный иммунитет
6.1. Митохондриальный противовирусный иммунитет
Клетки отвечают на вирусную атаку, активируя различные пути передачи сигнала, ведущие к продукции интерферонов [217], которые ограничивают или устраняют вторжение вируса. Присутствие вирусов внутри клетки сначала определяется рецепторами распознавания образов (PRR), которые распознают молекулярные образцы, связанные с патогенами (PAMP). PRR включают toll-подобные рецепторы (TLR), рецепторы, подобные домену олигомеризации нуклеотидов (NOD) (NLR), и рецепторы, подобные рецепторам (RLR), индуцируемым ретиноевой кислотой, ген I (RIG-I).Митохондрии были связаны с RLR, которые включают индуцируемый ретиноевой кислотой ген I (RIG-I) [218] и ген 5, связанный с дифференцировкой меланомы (Mda-5) [219]. Обе представляют собой расположенные в цитоплазме РНК-геликазы, распознающие дцРНК. N-конец RIG-1 имеет домены активации и рекрутирования каспаз (CARD), тогда как C-конец обладает активностью РНК-геликазы [218], которая распознает и связывается с не кэпированной и немодифицированной РНК, генерируемой вирусными полимеразами, зависимым от АТФазой образом. Это вызывает конформационные изменения и выставляет его CARD-домены для связывания и активации нижестоящих эффекторов, что приводит к образованию энхансосомы [220], запускающей продукцию NF κ B.RLR недавно были подробно рассмотрены [221–223].
CARD домен, содержащий белок, названный митохондриальной противовирусной передачей сигналов (MAVS) [15, 224], вирус-индуцированный сигнальный адаптер (VISA) [225], стимулятор промотора 1 IFN- β (IPS-1) [226] или Адаптер CARD, индуцирующий белок IFN- β (CARDIF) [227], действует ниже RIG-I. Помимо присутствия N-концевого домена CARD, MAVS содержит богатую пролином область и C-концевую гидрофобную трансмембранную (TM) область, которая направляет белок на внешнюю мембрану митохондрий и имеет решающее значение для его активности [15].ТМ-область MAVS напоминает ТМ-домены многих С-концевых заякоренных в хвосте белков на внешней мембране митохондрий, включая Bcl-2 и Bcl-xL [15]. Недавние сообщения показывают, что MAVS играет важную роль в индукции противовирусной защиты в клетке. Сверхэкспрессия MAVS приводит к активации NF κ B и IRF-3, что приводит к индукции интерферонового ответа типа I, который исчезает в отсутствие MAVS [15], что указывает на специфическую роль MAVS в индукции противовирусного ответа. .Также было показано, что MAVS предотвращает апоптоз за счет взаимодействия с VDAC [228] и предотвращения открытия MPTP.
6.2. Регуляция вирусами
Некоторые вирусы индуцируют отщепление MAV от внешних мембран митохондрий [227, 229], что значительно снижает их способность вызывать интерфероновый ответ. ВГС сохраняется в организме хозяина за счет снижения иммунного ответа клетки-хозяина, включая ингибирование продукции IFN- β посредством пути RIG-I [230–232]. Белок NS3 / 4A HCV колокализуется с митохондриальным MAVS [227, 229], что приводит к расщеплению MAVS по аминокислоте 508.Поскольку свободная форма MAVS не функционирует, удаление MAV из митохондрий инактивирует MAVS [227], что помогает парализовать защиту хозяина от HCV. Интересно, что другой член семейства Flaviviridae GB вирус B имеет 28% гомологию аминокислот с HCV по длине их открытых рамок считывания [233]. Белок NS3 / 4A вируса GB также расщепляет MAVS аналогично HCV, тем самым эффективно нарушая иммунный ответ хозяина, предотвращая продукцию интерферонов [234].Другие вирусы, такие как грипп A, перемещают компоненты RIG-I / MAVS в митохондрии инфицированных первичных макрофагов человека и регулируют противовирусные / апоптотические сигналы, повышая выживаемость вируса [235].
7. Вирусы захватывают митохондриальные белки хозяина
За прошедшие годы вирусы усовершенствовали различные стратегии для установления сложных отношений со своим хозяином с единственной целью — сохранить свое существование. Одна из таких стратегий включает захват митохондриальных белков клетки-хозяина.P32, клеточный белок, связанный с митохондриями, является членом комплекса, участвующего в импорте цитозольных белков в ядро. При входе в клетку аденовирус захватывает этот белок и использует его для транспортировки своего генома в ядро [236], тем самым увеличивая свои шансы на выживание и закрепление в клетке-хозяине. Во время сборки ВИЧ-1 изоакцепторы избирательно включаются в вирионы, связываются с геномом ВИЧ и используются в качестве праймера для обратной транскрипции [237].У человека единственный ген продуцирует цитоплазматические и митохондриальные Lys-тРНК-синтетазы (LysRS) путем альтернативного сплайсинга [238]. Митохондриальный LysRS продуцируется в виде препротеина, который транспортируется в митохондрии. Премитохондриальный или митохондриальный LysRS специфически упакован в ВИЧ [239] и действует как праймер для инициации репликации генома РНК ВИЧ-I, который затем связывается с сайтом, комплементарным 3′-концу 18 нуклеотидов. Предполагается, что вирусный белок ВИЧ R (Vpr) изменяет проницаемость митохондрий [138], что приводит к высвобождению премито- или мито-LysRS, который затем взаимодействует с Vpr [240] и упаковывается в вирионы потомства.
Виперин, белок, индуцируемый интерфероном, индуцируется в клетках в ответ на вирусную инфекцию [241]. Было показано, что этот белок предотвращает высвобождение частиц вируса гриппа из клеток, улавливая их липидными рафтами внутри клеток, тем самым предотвращая его распространение [242]. Во время инфекции HCMV индуцирует независимую от IFN экспрессию виперина, который взаимодействует с кодируемым HCMV белком vMIA, что приводит к перемещению виперина из ER в митохондрии. В митохондриях виперин взаимодействует с трифункциональным белком митохондрий и снижает выработку АТФ, нарушая окисление жирных кислот, что приводит к нарушению актинового цитоскелета клеток и усилению вирусной инфекционности [243].
8. Вирусы изменяют внутриклеточное распределение митохондрий
Вирусы изменяют внутриклеточное распределение митохондрий либо путем концентрации митохондрий рядом с вирусными фабриками, чтобы удовлетворить потребности в энергии во время репликации вируса, либо путем оцепления митохондрий в цитоплазме для предотвращения высвобождения медиаторов апоптоз. Белок X HBV вызывает опосредованное микротрубочками перинуклеарное кластеризацию митохондрий за счет активности динеина, опосредованной митоген-активируемой протеинкиназой p38 (MAPK) [244].Неструктурный белок 4A (NS4A) ВГС либо отдельно, либо вместе с NS3 (в форме полипротеина NS3 / 4A) накапливается в митохондриях и изменяет их внутриклеточное распределение [245]. Инфекция ВИЧ-1 вызывает кластеризацию митохондрий в инфицированных клетках [246]. Интересно, что ASFV вызывает опосредованное микротрубочками кластеризацию митохондрий вокруг вирусных фабрик в клетке, обеспечивая энергию для высвобождения вируса [247]. Подобные изменения наблюдались в фибробластах куриных эмбрионов, инфицированных вирусом лягушки 3, где были обнаружены вырожденные митохондрии, окружающие вирусные фабрики [248].
9. Вирусы имитируют митохондриальные белки хозяина
Молекулярная мимикрия — это «теоретическая возможность того, что сходства последовательностей чужеродных и собственных пептидов достаточно для перекрестной активации аутореактивных Т- или В-клеток пептидами, полученными из патогенов» [ 249, 250]. Поскольку структура следует за функцией, вирусы во время их совместной эволюции с хозяевами эволюционировали, чтобы имитировать белки-хозяева, чтобы достигать своих целей в ходе своего жизненного цикла внутри клетки.Имитация помогает вирусам получить доступ к клеточным машинам хозяина и значительно помогает им выживать во враждебной среде хозяина.
Mimivirus, член недавно созданного семейства вирусов Mimiviridae , кодирует эукариотический белок-носитель митохондрий (VMC-I) [251], который имитирует митохондриальный белок-носитель клетки-хозяина и, таким образом, контролирует транспортный механизм митохондрий в инфицированных клетках. Он помогает транспортировать ADP, dADP, TTP, dTTP и UTP в обмен на dATP, таким образом используя хозяина для удовлетворения потребностей в энергии во время репликации его A + T-богатого генома [251].Помимо VMC-I, мимивирус кодирует несколько других белков (L359, L572, R776, R596, R740, R824 L81, R151, R900 и L908) с предполагаемыми сигналами локализации митохондрий, которые предполагают, что мимивирус разработал стратегию захвата хозяина. митохондрии и использовали их физиологию для компенсации своих энергетических потребностей и биогенеза [251]. Вирусные гомологи Bcl-2 (vBcl-2) представляют собой другие группы вирусных белков, которые имитируют Bcl-2s клетки-хозяина и были описаны в другом месте этого обзора.
10.Вирусы вызывают истощение митохондриальной ДНК хозяина
Митохондрии млекопитающих содержат небольшой кольцевой геном, который синтезирует ферменты окислительного фосфорилирования и митохондриальные РНК (мтРНК) [27]. Чтобы увеличить шанс выживания, некоторые вирусы, по-видимому, приняли стратегию повреждения митохондриальной ДНК клетки-хозяина. Поскольку митохондрии действуют как источник энергии и играют важную роль в противовирусном иммунитете, возможно, что повреждение митохондриальной ДНК может помочь избежать митохондриальных противовирусных иммунных ответов [252].
Во время продуктивного инфицирования клеток млекопитающих in vitro , HSV-1 вызывает быструю и полную деградацию митохондриальной ДНК хозяина [252]. Белок UL12.5 HSV-1 локализуется в митохондриях и вызывает истощение ДНК в отсутствие продуктов других вирусных генов [252, 253]. Непосредственно ранний Zta-белок EBV взаимодействует с митохондриальным одноцепочечным ДНК-связывающим белком, что приводит к снижению репликации митохондриальной ДНК (мтДНК) и усилению репликации вирусной ДНК [254].HCV вызывает повреждение ДНК, опосредованное реактивными формами кислорода и закисью азота, в мтДНК хозяина [107, 255]. Интересно, что истощение мтДНК также наблюдалось у людей с коинфекцией ВИЧ / ВГС [256].
11. Выводы
Хотя был достигнут прогресс в понимании взаимодействия вирусов с митохондриально-опосредованными путями, пути, связывающие обнаружение вирусной инфекции с помощью PRR (или точный механизм, с помощью которого PRR распознают PAMP) и их связь с митохондриями -опосредованная гибель клеток остается малоизученной.Роль митохондрий в иммунитете и вирусных механизмах уклонения от них подчеркивает тот факт, что даже после миллиардов лет совместной эволюции борьба за выживание все еще продолжается. И хозяин, и вирусы развиваются, находя новые способы выжить. Может быть интересно отметить, что апоптоз, опосредованный митохондриями, мог бы быть эволюционной адаптацией, с помощью которой они могли бы эффективно предотвращать проникновение других микроорганизмов, пытающихся проникнуть в клетку-хозяина и, таким образом, эффективно утвердившись в качестве неотъемлемой части клетки.